| It
has been proposed that at high antigen concentrations lymphocytes bearing specific antigen
receptor sites can be destroyed by a mechanism involving complement (1-3). The same cells
can be stimulated to proliferate and secrete antibody at lower antigen concentrations.
Thus for a pure clone of cells, each with receptor sites of the same distribution and
specificity, antigen dose-response curves should show an ascending limb reflecting cell
stimulation at low antigen concentrations, and a descending limb reflecting cell
destruction at high antigen concentrations. If complement were removed from the system the
descending limb would be eliminated and the curves would plateau at high antigen
concentrations. Such ideal curves have been obtained in studies with lymph-node cells
cultured with the non-specific "antigen analogue"
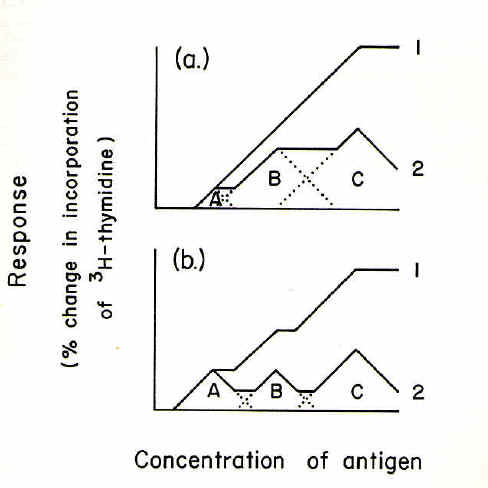
concanavalin-A in the presence and absence of complement (4,5). However, lymph-node cells are thought to consist of multiple clones each of a particular specificity for a given antigen (1,3,6). Thus the pattern of dose-response curves with lymph-node cells would be expected to differ from the ideal. This was found in studies of the stimulation by specific antigen of the radioactive labelling with [3H]-thymidine of populations of lymph-node cells from immunized rabbits (7). Dose-response curves for cultures containing autologous serum preheated at 37oC ("37oC-serum") diverged below the curves for cultures containing 56oC-serum. It was shown that this pattern of dose-response curves could be interpreted in terms of a successive recruitment of low specificity cells with increasing concentrations of antigen. The interpretation is given in Fig. 1a which shows theoretical antigen dose-response curves for three sub-populations (clones) of cells (A, B, C) bearing receptors of decreasing specificity for antigen. As the concentration of antigen increases, clone A begins to be inhibited but clone B begins to be stimulated. At still higher antigen concentrations clone B begins to be inhibited but clone C begins to be stimulated. The total response at any particular antigen concentration reflects the difference between the stimulatory and inhibitory effects of that antigen concentration. The response over the range of antigen concentrations is shown in curve 2. If the complement-dependent inhibitory component is removed, curve 1 is generated.
To examine this interpretation, rabbits were immunized according to either (i) a "standard" protocol (1 mg of keyhole limpet hemocyanin, KLH, 2 weeks before culturing popliteal lymph-node cells), or (ii) a protocol designed to increase the proportion of high specificity cells (0.1 mg of KLH 4 weeks before culturing cells). As shown in Fig. 1b, it was predicted that the divergence between the dose-response curves would be greater when the latter protocol was employed, particularly at low antigen concentrations. Ths was found to hold experimentally. In other experiments it was found that the percentage stimulation of [3H]-thymidine incorporation by KLH was enhanced by 2-mercaptoethanol (2-ME) to the same extent as the enhancement by 56oC-serum. In combination the two enhancements were mutually exclusive and non-additive suggesting that 56oC-serum and 2-ME were acting by a common mechanism. Antigen dose-response curves in 37oC-serum diverged below those for cultures containing 37oC-serum with 2-ME; this divergence, like the divergence with 56oC-serum, was greater in the case of cells from rabbits given the protocol designed to increase the proportion of high specificity cells.
REFERENCES 1. Burnet, F. M., in "The Clonal Selection Theory of Acquired Immunity," p. 64. Cambridge University Press, 1959. 2. Azar, M. M., J. Immunol. 106, 241 (1971). 3. Forsdyke, D.R., J. Theor. Biol. 52, 187 (1975). 4. Milthorp, P., and Forsdyke, D. R., Biochem. J. 132, 803 (1973). 5. Forsdyke, D. R. , Proc. Can. Fed. Biol. Socs. 19, 245 (1976). 6. Siskind, G. W., and Benacerraf, B., Adv. Immunol. 10, 1 (1969). 7. Forsdyke, D. R., Immunology 25, 597 (1973). 8. Soulillou, J. P., Carpenter, C. B., Lundin, A. P., and Strom, T. B., J. Immunol. 115, 1566 (1975). |