The " Residue " or " Base " upon which "Unit" or "Transferable" Characters are "Imposed" or "Implanted"
![]()
Report I to the Evolution Committee (1902)Report 2 to the Evolution Committee (1904)
Facts Limiting the Theory of Heredity (1907)
The Progress of Genetics (1907)
Mendel's Principles of Heredity (1909)
Heredity (1914)
Evolutionary Faith and Modern Doubts (1922)
Progress in Biology (1924)
Segregation (1926)
Distinction between Sporadic and Non-Sporadic Sterilities (Summary 2008)
The B in "BDM" (2010)
![]()
|
Thomas Huxley described himself as "Darwin's bulldog." Although Bateson did not use the term, Bateson is now widely recognized as "Mendel's bulldog." Less well-known is that Bateson was also "Romanes' bulldog," albeit cryptically. Whereas the Darwinians failed to understand Romanes' 1886 Linnean Society address on "Physiological Selection", the 26 year old Bateson immediately saw what Romanes was getting at, and wrote to his sister, who was then working with one of Darwin's sons: |
|
"I don't agree with you that Romanes' paper is poor. It seems a fair contribution and at all events does, as he says, put the whole view on a much more logical basis. The scheme thus put will at least work logically while the other, as left by Darwin, would not. ... I did not suppose Romanes would ever write as good a paper. ... it is a straight forward, common-sense suggestion." |
|
Another "Romanes bulldog" was Joseph T. Cunningham who chided Bateson for not acknowledging that the ideas he advocated were similar to those of Romanes. However, Bateson, who made his living as Steward of his college, needed an academic appointment. Romanes had put many backs up, including that of the Professor of Zoology and Comparative Anatomy, Alfred Newton (1829-1907). Why, we do not know, but in one of his books (1882) Romanes came down on the side of those who had publicly questioned Newton's case that cuckoos might colour their eggs to match those in the host nest. In a letter to John Gulick (Sept. 2 1888) Romanes wrote that Newton "hates me with a deadly hate." Bateson held a tenuous foothold in Newton's Department of Animal Morphology until 1899 when he became Newton's Deputy. Bateson and Newton were of different political persuasions (Liberal versus Tory) and did not see eye-to-eye on the issue of giving academic equality to women. It is possible that, had Bateson openly supported Romanes' views, Newton would have been further offended (for more Click Here). |
Report I to the Evolution Committee. Bateson & Saunders 1902
"Though some degree of sterility on crossing is only one of the diverse properties which may be associated with Specific difference [differentiation of species], the relation of such sterility to Mendelian phenomena must be a subject of careful enquiry. ...The sterility of certain crosses is merely an indication that they cannot divide up the [parental] characters among their gametes. If the parental characters, however dissimilar, can be split up [segregated], the gametes can be formed, and the inability to form gametes may mean that the process of resolution [separation of characters] cannot be carried out. ... Has a given organism a fixed number of unit-characters? Can we rightly conceive of the whole organism as composed of such unit characters, or is there some residue -- a basis -- upon which the unit characters are imposed?... We are thus brought to face the further question of the bearing of Mendelian facts on the nature of species. The conception of Species, however we may formulate it, can hardly be supposed to attach to allelomorphic or analytical varieties [characters associated with conventional Mendelian genes]. We may be driven to conceive "Species" as a phenomenon belonging to that "residue"... ." |
![]()
Report 2. Reports to the Evolution Committee of the Royal Society.
By W. Bateson, E. R. Saunders & R. C. Punnett (1904) p. 92.
"[Withered] anthers were seen from time to time in many families, though commonly confined to individual flowers. This sporadic sterility has not been particularly studied. It is of interest to compare this example of the definite appearance of sterility ... with the familiar [regular] occurrence of sterility in cross-breds [hybrids]. Such a phenomenon has often been supposed to indicate remoteness of kinship, yet here a closely comparable effect occurs ... as the result of a cross between two types which must be nearly related. Mr. Gregory in a careful examination of the pollen-genesis found that the divisions were normal up to the reduction division, when the chromosomes formed shapeless knots and entanglements, failing to divide." |
![]()
Facts Limiting the Theory of Heredity (1907) Science 26, 649-660.
"While knowledge of late progressed so rapidly in regard to many genetic phenomena, we still know next to nothing of the facts relating to the incidence of partial sterility among heterozygous forms. Guyer found that the abnormality of which the sterility of hybrid pigeons is the expression, begins in the reduction division [of meiosis] and is apparent as an entanglement of the chromosomes which fail to divide. In many cases sterility is partial; and, for example, a proportion of good pollen grains occurs mixed with aborted grains." |
![]()
The Progress of Genetics Since the Rediscovery of Mendel's Papers
(1907) Progressus rei Botannicae 1, 368-418.
| "Cytological appearances: sterility
...the relation of Mendelian observations to the facts of cell division. Nevertheless it should be remarked that all that has been observed by cytologists is consistent with the results of experimental genetics. The recognition of a definite differentiation among the chromosomes (see especially Sutton [1902; Biol. Bull. 4, 24]) is probably an important advance, though until we can positively recognize characters in the zygote as associated with some visible cytological elements we must be cautious in forming positive conclusions as to the relation of cytological appearances to the phenomena of heredity. Such an observation has however been lately made by E. B. Wilson ([1906; J. Exp. Zoology 3, no 1]) who has found evidence of a visible dimorphism among the sperm cells of certain insects, and has shown that this dimorphism corresponds with differences existing between the somatic cells of the males and females of these species. ... There is still no positive proof that segregation occurs at the reduction division, but all the facts known point to that conclusion. Indeed we can scarcely doubt that this is the critical moment. Investigations of sterile forms (Guyer [Chicago 1990 dissertation]; Gregory [1905. Proc. Camb. Phil. Soc. 13, 148]) show that it is often at this division that abnormality begins. Nevertheless I am not yet convinced that segregation is exclusively confined to that division ... in the somatic cell divisions similar processes may be concerned. That segregation precisely similar to that which occurs among gametes may take place in somatic divisions is indeed proved by the existence of bud-sports. The appearance of a glabrous Nectarine on a Peach tree or of a white flower on a red-flowered plant is unquestionably due to segregation, and it is interesting to observe that the characters which segregate in these "sports" are often those which we know to segregate in gametogenesis. Such facts raise the fundamental question whether in the normal process of somatic differentiation, segregations of a similar nature may not occur ... . [includes phenomena now known as "loss of heterozygosity" or "gain of homozygosity" due to gene conversion (non-reciprocal recombination)]
The Relation of Mendelian Segregation to Specific [Species] Distinctions. Mendelian segregation proves the unity of characters. Is it a sound deduction that specific differences [differences fundamental to the differentiation of species] come into existence by the addition or elimination of such character units? ... It is scarcely necessary to insist that plenty of the characters which are now known to segregate would be far more than sufficient to constitute specific differences in the eyes of most systematists ... . But this is not enough. We must eventually go further; and ... we must ask whether the results of ... investigation will show
We may indeed, as I have said elsewhere ([Reports to the Evolution Committee. 1902]), be driven to conceive specific difference as a property of the residue or basis upon which the allelomorphic characters are implanted; but it is not easy to suppose that the features, breadth of leaves, and length of flowering stem -- named by de Vries as non-splitting [segregating] characters in Lychnis -- are of this fundamental nature. ... [dated] August 1906."
[For one way of looking at implanting on a base or residue, see the figures of Goldschmidt (1917) as reproduced by Richmond and Dietrich (2002; Genetics 477-482.] |
![]()
Mendel's Principles of Heredity (1909)
"Possible
Limits to Recombination.
|
![]()
(Full text of 1914 Address proposing genetic information as a 'phenomenon of arrangement', which we can now interpret as a sequence of bases in DNA.) (Click Here)
![]()
Evolutionary Faith and Modern Doubts
By W. Bateson (1922) Science 55, 55-61 and Nature 109, 553-556.
Bateson frequently refers to the work of Michael F. Guyer (1874-1959; see Guyer webpage (Click Here)). Bungener and Buscaglia (2003; Hist. Phil. Life. Sci 25, 27-50) reveal that there is material in Guyer's 1900 thesis that is not in the 1902 paper. In particular, they note from page 51 of the thesis that Guyer:
|
![]()
An interest of the following interchange between Bateson and his critic C. R. Crowther, is the "sword and scabbard" analogy, which can now be extrapolated to the stereospecific pairing between DNA bases in homologous chromosomes. Crowther points out that the requirements for cooperation, which, if it failed, would produce a inviable hybrid, should be less stringent than the requirements for meoitic pairing ("conjugation"), which, if it failed, would produce a sterile hybrid. Crowther imagines that the latter failure, due to differences between the homologs, would be the result of: "a gradual modification, spread over a length of time, and involving many chromosomes". |
|
|
![]()
By W. Bateson (1924)
From William Bateson FRS, Naturalist. His Essays and Addresses. Cambridge University Press, 1928. p. 406-408.
"If we now have to recognize that the transferable [genic] characters do not culminate in specific distinctions [cause the origin of species], the acknowledgement will not come from us alone. The old belief of systematists that real species differ from each other in some way not attainable by summation of varietal [genic] characters is no longer contestable, and we know now upon what to concentrate. It is no occasion for dismay. We have not to go back very far. We do not understand specific differences [speciation], nor can we account for the adaptive mechanisms. Was it to be expected that we should? Biology is scarcely a century old, and its intensive study is of yesterday. There is plenty of time ahead.
The
identification of the transferable characters and their linkages has led to a further
discovery of the greatest ... brilliancy, which must have consequences as yet
unestimable.
Morgan and his colleagues have ... proved that some, probably all, of
this group of characters are determined by elements
transmitted in or attached to the chromosomes....
|
![]()
J. Genetics 16, 201-235 (1926)
"In 1922 I had the honour of delivering the Leidy Memorial Lecture at Philadelphia. The substance of that address is embodied in the following paper. I was reluctant to publish it immediately for several reasons. The evidence to which I appealed had not been fully tested. Some also might be disposed to set aside the phenomena adduced, on the ground that they are exceptional and of small immediate significance pending the determination of broader principles. Experience has meanwhile provided confirmation where it was required, and several of the classes of segregation which might formerly be held peculiar or anomolous, have been demonstrated in such abundance that no survey of genetical physiology can disregard them. In so far, moreover, as I was concerned with principle, I wished for opportunities of hearing the criticism of other geneticists holding different opinions; but after such debates I have learned nothing which appears to dispose of the views to which I inclined.
... Having in view the various facts and considerations here enumerated, I think we shall do genetical science no disservice if we postpone acceptance of the chromosome theory in its many extensions and implications. Let us distinguish fact from hypothesis. It has been proved that, especially in animals, certain transferable [genic] characters have a direct association with particular chromosomes. Though made in a restricted field this is a very extraordinary and most encouraging advance. Nevertheless, the hope that it may be safely extended into a comprehensive theory of heredity seems to me ill-founded, and I can scarcely suppose that on a wide survey of genetical facts, especially those so commonly witnessed among plants, such an expectation would be entertained. For the phenomena to which the simple chromosome theory is inapplicable, save by the invocation of a train of subordinate hypotheses, have been there met with continually, as even our brief experience of some fifteen years has abundantly demonstrated. Through all this work, with ever increasing certainty, the conviction has grown that the problem of heredity and variation is intimately connected with that of somatic differentiation, and that in an analysis of the interrelations of the two manifestations of cellular diversity lies the best prospect of success. Pending that analysis, the chromosome theory, though providing much that is certainly true and of immense value, has fallen short of the essential discovery." |
![]()
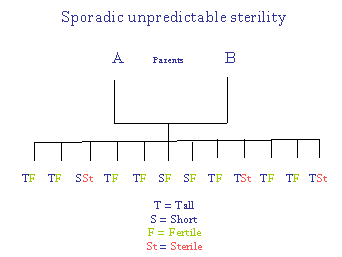
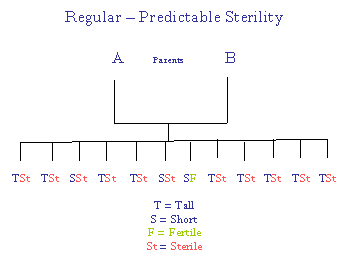
Distinction between Sporadic and Non-Sporadic Sterilities
|
|
For sporadic sterility Mendelian (3:1) ratios are followed. For non-sporadic sterility, all (or most) offspring are sterile.
|
|
|
Thus, sporadic sterility referred to genes, whereas non-sporadic sterility referred to the "base" or "residue" upon which the genes were considered to be implanted. It is proposed by the author of these pages that the residue can be equated with the "accent" of DNA - the percentage (G + C) in DNA. For more, please see:
Two Levels of Information (1999) Romanes, Bateson & Darwin's "Weak Point" (2010) The B in "BDM" (2010)
|
![]()
Go to: Bateson and Saunders, 1902 (Click Here)
Go to: Heredity & Variation, 1909 (Click Here)
Go to: Bateson's Melbourne Address 1914 (Click Here)
Go to: Bateson on the "Base" or "Residue" (Click Here)
Go to: Opposition to Bateson (Click Here)
Go to: Bateson & Goldschmidt (Click Here)
Go to: Video Lecture on Bateson & Romanes (Click Here)
Return to: Evolution Index (Click Here)
Return to: Home Page (Click Here)
![]()
This page was established circa 2000 and last edited 12 January 2014 by Donald Forsdyke