|
AIDS
CURE
TREATMENT BY PROGRAMMED LYMPHOCYTE ACTIVATION
Inductive Therapy to "Flush Out"
Latent Viruses, Plus Conventional
Therapy to Strike "On-the-Wing"!
to Strike "On-the-Wing"!
Pangens, Rinderpest, and Virus
Latency (1868)
Grouse Shooting and Antibiotic Resistance
Programmed Activation of
T-Lymphocytes (1991)
Quotations from Chun et
al. (1998)
Quotations from Ho (1998)
Attack Reservoirs (Steve Bunk
1998)
COMMENTARY (The Scientist, Jan 2000)
Definition of "Antibiotic"
The Truth Dawns
Bioinformatic analysis of
potential target sites (1995)
Codon Choice in Retroviruses
Death While Seeking the Origin of
AIDS
More of the Same in South Africa
The Drug Companies Win the "Image Game"
HIV Recombination and Speciation (2013)

Pangens, Ringerpest, and
Virus Latency
In developing his hypothesis of "pangenesis",
Darwin (1868) was encouraged by
reports (Romano,
1997, 2002) of the great proliferative powers of microorganisms,
as related in the 1866 Third Report of the Commissioners
on the Cattle
Plague, Rinderpest, which is now known to be caused by a Morbillivirus
(Click Here):
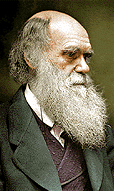 |
Nor
does the extreme minuteness of the gemmules [small
"germs"], which can hardly differ much in
nature from the lowest and simpliest organisms, render it improbable that
they should grow and multiply. A great authority, Dr. Beale, says:
"that minute yeast
cells are capable of throwing off buds or gemmules, much less than the 1/100000
of an inch in diameter;" and these he thinks are
"capable of
subdivision practically ad infinitum". |
|
A
particle of small-pox matter, so simple as to be borne by the wind, must multiply itself
many thousandfold in a person thus inoculated; and so [ also is the
case] with the
contagious matter of scarlet fever. It has recently been ascertained that a minute portion
of the mucous discharge from an animal affected with rinderpest, if placed in the blood of
a healthy ox, increases so fast that in a short space of time
'the whole mass of blood, weighing many pounds,
is infected, and every small particle of that blood contains enough poison to give, within
less than forty-eight hours, the disease to another animal.'"
|
The hypothesis of pangenesis required modification by Hugo de Vries (1889) to make it approximate to modern concepts
(e.g. the "Weismann
barrier" that prevents transmission of genes from soma to the germ-line).
Darwin's point
was reiterated in 1922 by H. J. Muller when commenting on observations by Canadian Felix
d'Herelle on agents (viruses) which infect bacteria (now known as bacteriophages):
"That two distinct kinds of substances - the
d'Herelle substances and the genes - should both possess this most remarkable property of
heritable variation or 'mutability', each working by a totally different mechanism, is
quite conceivable, considering the complexity of the protoplasm; yet it would seem a
curious coincidence indeed. It would open up the possibility of two totally different
kinds of life, working by different mechanisms.
On the other hand, if these
d'Herelle bodies were really genes, fundamentally like our chromosomal genes, they would
give us an utterly new angle from which to attack the gene problem. They are filterable,
to some extent isolatable, can be handled in test-tubes, and their properties, as shown by
their effects on bacteria, can then be studied after treatment.
It would be very rash to call
these bodies genes, and yet at present we must confess that there is no distinction known
between genes and them. Hence we cannot categorically deny that perhaps we may be able to
grind genes in a mortar and cook them in a beaker after all. Must we geneticists become
bacteriologists, physiological chemists, and physicists, simultaneously with being
zoologists and botanists? Let us hope so." |
Darwin's idea that "gemmules" could transfer from one cell
to another, where they could become part of the genetic material of the new cell, finds a
modern analogy in the phenomenon of viral latency, where viral nucleic acid
seamlessly integrates and
hides in the genome of its host. In the case of HIV, the virus is not known to enter the
germ line.
Nevertheless, our genomes are littered with retroviral remnants, indicating
that, in the past, HIV-like viruses, similar to Darwin's proposed "gemmules,"
have transferred somatically-acquired information to the germ cells, and hence to the
offspring. Gregory Bateson (1979)
considered that "gemmules"
in the form of RNA molecules, might transfer information from a blacksmith's
biceps to his germ-line (hence
facilitating the Lamarckian inheritance of acquired characters),
and questioned the absolute nature of the Weismann barrier:
| "But that assumption does not look
so safe today as it did twenty years ago. If RNA can carry imprints of
portions of DNA to other parts of the cell and possibly to other parts
of the body, then it is imaginable that imprints of chemical
changes in the biceps could be carried to the germ-plasm." |
Indeed, we now
know that RNA can spread information systemically throughout plant tissues (Waterhouse
et al. 2001), but there
is no evidence that the spread can include germ-line tissues (albeit
the distinction between germ-line and soma in plants in less distinct than in
animals). In
microorganisms the "memory" of prior infection is transferred to
descendants in the CRISPR system so this, in the words of Koonin and Wolf (2009),
would indeed constitute "a bona fide Lamarckian mechanism."
Darwin,
C. (1868) The Variation
of Animals and Plants under Domestication. Chapter 27. Murray, London.
Koonin, E. V. & Wolf Y. I. (2009) Is
evolution Darwinian or/and Lamarckian? Biology
Direct 4:42
Liu, Y.
(2006) Advances
in Genetics 56,
101-129. Historical and modern genetics of plant graft hybridization.
Muller,
H. J. (1921) American Naturalist 56, 32-50.
Bateson,
G. (1979) Mind
and Nature. Dutton, N. Y. p.
166.
Romano,
T. M. (1997) The cattle plague of 1865 and the reception of "the germ
theory" in mid-Victorian Britain. J.
Hist. Med. Allied Sci. 52,
51-80.
Romano, T. M.
(2002)
Making Medicine Scientific. John Burdon Sanderson and the
Culture of Victorian Science.
John Hopkins University Press.
Third Report of The Commissioners appointed to inquire
into The Origin and Nature, etc. of The Cattle Plague. (1866) House of
Parliament, London (Click Here)
Vries, H. de
(1889) Intracellular Pangenesis. Fischer, Jena.
Waterhouse,
P.M., Wang, M-B., & Lough, T. (2001) Gene silencing as an adaptive
defence against viruses. Nature
411,
834-841. |
Grouse Shooting and
Antibiotic Resistance
A proposed treatment of AIDS, taking into account the phenomenon
of viral latency, only began to receive serious attention several years after its formal
presentation in 1991. Its underlying principle: |
The
fundamental tenet of antibiotic usage is that, wherever possible, one hits the pathogen
with a dose of drug sufficient to kill all
organisms in a short period, thus not allowing antibiotic resistant strains to emerge. |
HIV has the latency option which allows it to "sit out" until the antiviral antibiotic gunfire
has subsided. Biomedical researchers have long known this, yet they have continued to hunt for more and more antiviral antibiotics that they
knew could only be
administered under conditions (i.e. long-term therapy) such that resistance in the
pathogen would be actively fostered. |
|
Thus some of the most effective antiviral
agents against HIV have been rendered useless.
|
Had research funds been
available to those researchers who painfully spelled out, time and time again, in grant
application after grant application, the overwhelming importance of investigating virus latency, we might have avoided the
antibiotic resistance problem and by now had an effective, cheap, short term, cure for
AIDS.
In principle, the approach is quite
simple, as a grouse shooting metaphor can show. In the grouse shooting season the woods
and copses echo not only gun-fire, but the thwack of the beaters. To rid land of
grouse requires a two-fold approach:
- (i) Beaters to get the birds to fly
up.
- (ii) Guns and good shooters.
The beaters alone
will just cause the grouse to spread to other sites.
The shooters alone will just be able to shoot the occasional
grouse which is so unfortunate as to expose itself.
 The combination is lethal! The combination is lethal! |
In the AIDS context, the
guns are drugs such as AZT, and a complex of drugs including inhibitors of a
viral protease ("combination therapy" or "HAART"). These drugs hit AIDS viruses "on the wing", but are useless
against latent virus which hides, usually in DNA form, integrated into the DNA of
its host cell. We need drugs to simulate the beaters.
In 1991 it was suggested that cytokines
such as TNF-alpha might fill this role. It was not until 1998 that major laboratories in
the field began to recognize this, although the most influential still
believed that the emergence of resistant strains was "inevitable".
But by 2003 work from Dean Hamer's laboratory had shown:
"Following
reactivation viral expression was dramatically increased,
rendering the infected cells susceptible to an anti-HIV
immunotoxin. Treatment with the immunotoxin in conjunction
with agents that activate virus expression without
inducing cell division (IL-7 or the non-tumor-promoting phorbol
ester prostratin) depleted the bulk of the latent reservoir and
left uninfected cells able to respond to subsequent costimulation.
We demonstrate that activation of latent virus is required
for targeting by antiviral agents and provide a basis for
future therapeutic strategies to eradicate the latent
reservoir." |
The strategy even acquired its own acronym
"IAT" (immune activation therapy; Kulkosky & Pomerantz 2002).
Similar progress was achieved by Jerome Zack and his colleagues. Even the New
York Times (Sept.
23rd, 2003) began to sit up:
"In
the U.C.L.A. study, released last week by the journal Immunity,
Dr. Zack described choosing his drugs - prostratin and
interleukin-7, to "tickle the cell, just turn the virus on
without turning on the cell." |
While perhaps unduly cautious about not
turning on the cell (which would be destroyed by its own activated virus)
in the paper, Brooks et al. noted:
"Our
studies demonstrate in primary cells the functional dormancy of
HIV in the latent state and consequently, the necessity for viral
reactivation for recognition by antiviral agents." |
They also noted that in latent HIV there
is a low level of virus RNA production that is aborted. They speculate
that:
"Latency
is not necessarily maintained by active inhibition of the LTR, but
instead ... there is a lack of specific cellular proteins in
quiescent T cells that are necessary for productive viral
expression. Exactly which factors these are is currently unclear,
but in their absence the virus is dormant. ...These cellular
transcription factors must be induced early following stimulation
and prior to a substantial increase in cellular RNA
production." |
Candidates for such transcription
factors would include G0S30/EGR1 and other putative "G0/G1 switch genes"
Click Here.
Some of the pathways involved in T
cell activation involve "nuclear factor kappa B" (NFkB; see
below), which can be activated by agents reacting with a particular
receptor at the cell surface known as "Toll-like receptor 5" (TLR)
- present on T cells (Williams et al. 2004; Caron et al. 2005). In 2008
Burdelya and coworkers showed that a bacterial polypeptide (derived from
Salmonella flagellin and code named CBLB502) could activate TLR5, but the
possibility of its use in AIDS therapy was not even mentioned. In
2009 Richman and coworkers renewed the appeal for IAT, pointing out the
long-term cumulative toxicities of HAART and foreseeing an era "in
which HAART is no longer a lifetime necessity."
-
Berger, E. A., Moss, B. & Pastan, I. (1998) Reconsidering target toxins to
eliminate HIV infection: You gotta have HAART. Proc.
Natl. Acad. Sci. USA 95, 11511-11513.
- Bocklandt, S., Blumberg, P. M. & Hamer, D. H.
(2003) Activation of latent HIV-1 expression by the potent anti-tumor
promotor 12-deoxyphorbol 13-phenacetate. Antiviral
Research 59, 89-98.
- Brooks, D. G., Hamer, D. H., Arlen, P. A., Gao, L.,
Bristol, G., Kitchen, C. M. R., Berger, E. A. & Zack, J. A. (2003)
Molecular characterization, reactivation and depletion of latent HIV. Immunity
19, 413-423.
- Burdelya, L. G. et al. (2008) An agonist of Toll-like receptor
5 has radioprotective activity in mouse and primate models. Science
320, 226-230.
- Caron, G. et al. (2005) Direct stimulation of human T cells via
TLR5 and TLR7/8: flagelling and R-848 up-regulate proliferation and IFN-gamma
production by memory CD4+ T cells. J.
Immunol. 175, 1551-1557.
- Chun et al., (1998). Induction of HIV-1 replication in latently infected CD4 T
cells using a combination of cytokines. J. Exp. Med.
188, 83-91.
- Cohen, O. J. & Fauci, A. S. (1998) HIV/AIDS in 1998 -
Gaining the upper hand? J. Amer. Med. Assoc.
280, 87-88.
- Cohen, O. J. & Fauci, A. S.(1999) Transmission of
drug-resistant strains of HIV-1: unfortunate, but
inevitable. Lancet
354, 697-698.
- Forsdyke, D. R. (1991). Programmed activation of T-lymphocytes. A theoretical
basis for short term treatment of AIDS with Azidothymidine. Medical
Hypothesis 34, 24-27.
- Ho, D. D. (1998). Towards HIV eradication or remission. Science 280, 1866-1967.
- Kulkosky, J. & Pomerantz, R. J. (2002) Approaching
eradication of highly infectious antiretroviral therapy - persistent
HIV-1 reservoirs with immune activation therapy. Clinical
Infectious Diseases 35, 1520-26.
- Maeda, M. et al. (2006) Tristetraproline [G0S24]
inhibits HIV-1 production by binding to genomic RNA. Microbes
and Infection 8, 2647-2656.
- Richman, D. D., Margolis, D. M., Delaney, M., Greene,
W. C., Hazuda, D. & Pomerantz, R. J. (2009) The challenge of finding
a cure for HIV infection. Science
323, 1304-7.
- Williams, S. A., Chen, L-F., Kwon, H., Fenard, D.,
Bisgrove, D., Verdin, E. & Greene, W. C. (2004) Prostratin
antagonizes HIV latency by activating NF-kB. J.
Biol. Chem. 279, 42008-17.[This
paper cites Forsdyke 1991.]
|
 |
Programmed Activation of T-Lymphocytes.
A Theoretical Basis for Short Term Treatment of AIDS with Azidothymidine
D. R. Forsdyke. Medical Hypothesis (1991) 34, 24-27. (With copyright permission
from Academic Press)
Abstract
Introduction
Renewal
of the T-lymphocyte population does not require feed-back to the stem-cell level
HIV DNA is destroyed when
the host cell is destroyed
Mechanism of action of AZT
Need for programmed
synchronous activation of all host T cells
HIV infection of non-lymphoid tissues
Abstract - When its T-lymphocyte host cell is activated,
the latent (DNA) form of human immunodeficiency virus (HIV) is activated to produce RNA
copies which are liberated as virus particles from the cell. In this process the cell is
destroyed together with the latent virus. If administered at this time, 3'-azidothymidine
(AZT) would specifically prevent the liberated RNA copies replicating and establishing
latency in new host cells. The RNA copies would then be degraded by viral or host
ribonucleases. Thus, one DNA copy of HIV and its RNA progeny would be eliminated from the
body.
However, many DNA copies of HIV would remain in other
cells ("latency"). The
main problem of therapy with AZT is that activation of host cells to become permissive for
production of virus is random
in time. Activation depends on chance encounters of an infected
person with the particular foreign antigens to which individual T-cells
bearing latent HIV can specifically respond. It is primarily for this reason that AZT must
be administered continuously.
If all T-cells
could be polyclonally stimulated at
one time, all
HIV-bearing T-cells would be destroyed and concomitant administration of
AZT for a short term
would prevent the replication of all
liberated viruses.
Unlike most renewable end cells in the body, the
maturation of T-cells involves processes of positive and negative
selection. To preserve the 'educated'
T-cell population, T-cell renewal occurs at the end cell
level, rather than at the stem cell level. It is possible that normal physiological signals concerned
with this homeostatic regulation of T-lymphocyte population size could be
harnessed to
produce synchronous
activation of all T-lymphocytes. Tumor necrosis factor-alpha
(TNF) has
some of the properties expected of a postulated
polyclonal activator needed for this programmed activation of T-lymphocytes. |
Introduction
Prolonged treatment with 3'-azidothymidine (AZT) extends the life
of patients with AIDS who are infected with the human immunodeficiency virus
(1). Recent
evidence suggests that asymptomatic HIV-seropositive individuals can also benefit from
prolonged AZT treatment (2). The continuing exponential increase in the number of
seropositive individuals and the need for prolonged treatment with this expensive drug
threatens to seriously burden healthcare systems worldwide.
To be
sure of curing AIDS, all forms of HIV within the body, both free and latent, need to be
eradicated. Although there are many recent reviews on AIDS treatment strategies
(2 - 6),
to our knowledge none of these considers the possibilities that there might be conditions
under which AZT could be employed:
We here consider various aspects of
lymphocyte biology, the HIV life-cycle, and the mechanism of action of AZT, which lead us
to a more optimistic assessment of the role of AZT and related drugs in the treatment of
AIDS. |
Renewal
of the T-lymphocyte population does not require feed-back to the stem-cell level
|
The
'education' of T-lymphocytes involves both
positive and negative selection (7, 8). Positive selection
generates sets of T-lymphocytes with the potential to respond to various
'self' determinants (e.g. MHC, CDI, TI and Qa-1 antigens; 9-11). Negative selection eliminates cells responding to self
with high specificity. The final immunological repertoire consists of numerous small
clones of cells. Members of a particular clone are each capable of recognizing a
particular set of 'nearself' antigenic determinants with
varying degrees of specificity.
The range
of specific responsiveness exhibited by an individual reflects the outcome of these
selection processes (and further positive selections by foreign antigens), over many
years. To renew the educated T-lymphocyte population after depletion
(perhaps due to haemorrhage), could be a protracted process if renewal required
reeducation. Individual T-cells (end cells), rather than stem cells,
should be responsive to homeostatic control mechanisms affecting the size of the total T-lymphocyte
population (7). Thus peripheral immunologically-competent clones of T-cells
should be responsive not only to the cues provided by foreign antigenic determinants
(through the determinant-specific T-cell antigen receptor) but also to
cues provided by the growth factors concerned with T-lymphocyte
population size homeostasis (through appropriate receptors). This self-renewing property
of the peripheral T-lymphocyte population is now established
experimentally (12-14). |
HIV DNA is destroyed when the host cell is destroyed
Like other retroviruses, HIV integrates into the DNA of its
host-cell and can remain there in quiescent form for prolonged periods (15,
16).
Activation of latent viruses generally requires concomitant activation of their host
cells, which then become permissive for virus production (17). In the case of HIV-infected
'resting' T-lymphocytes this activation
appears to require reaction with antigen or lectin (18). Subsequent intracellular signals
result in rapid changes in at least one protein encoded by a host gene (NFkB;
19). This
protein may then transmit activation signals to other host genes which play a role in the
switch from the resting (G0) phase to the activated (G1) phase of
the cell cycle and/or in progression through the G1 phase (20,
21). The protein
may also transactivate HIV genes (22,
23). The activated HIV genome (DNA) is then
transcribed to generate RNA copies of itself which are eventually packaged and released.
The host cell with its associated HIV DNA is destroyed in this process.
Thus cell
activation is therapeutic to the
extent that the latent DNA form of the virus is destroyed. The virus is triggered to
destroy itself. The problem, of course, is that newly liberated viruses in RNA form infect
new cells and can then establish latency in these cells. Effective therapy of AIDS
requires a drug, or drugs, which can achieve two,
preferably concomitant, results:
- The first of these is activation of all host cells
carrying latent virus so that such cells will be destroyed by the virus.
- The second is the prevention of liberated viruses
infecting, replicating in, and establishing latency in, previously uninfected cells
|
Mechanism
of action of AZT
The ideal therapeutic agent is a 'magic
bullet' which exploits some difference
between the metabolisms of a pathogen and its host. The replication of HIV is strictly
dependent on the viral enzyme reverse transcriptase, which is a DNA polymerase generating
DNA from the viral RNA template. This enzyme is widely distributed in both eukaryotes and
prokaryotes (24), but has not been shown to play a critical role in
eukaryotes. The enzyme
differs from the major host DNA polymerase activity in not being in a multienzyme complex
(25, 26), and not being capable of proof-reading DNA. The nucleotides which are linked
together linearly to make DNA are not scrutinized for abnormalities (27,
28). The
resulting high error rate results in a diversity of forms which may be advantageous for
the virus (29).
The
most widely accepted view of the selectivity of the action of the nucleotide analogue AZT
is that it is added by the viral reverse transcriptase to the elongating copy of viral
DNA, but is rejected by the proof-reading activity of host DNA polymerase. Thus
replication of HIV-RNA is selectively interrupted (3, 4,
30). Ribonuclease associated with
the reverse transcriptase, or a host ribonuclease, would probably then destroy the RNA so
that it could not act as a template for more copies of itself. |
Need for programmed synchronous activation of all host T cells
The switch of a G0 T-lymphocyte
containing HIV DNA in its genome to the G1 state permissive for viral
replication can occur under two circumstances:
The cell can be
activated by an antigenic determinant corresponding to the cell's specific
antigen-receptor (monoclonal response). The timing of this event would be entirely
dependent on random chance encounter of the infected person with that particular antigen.
Alternatively, the
cell can be activated as part of a polyclonal homeostatic response to a lectin-like factor
(lymphokine, growth factor) which signals a need to restore the size of the T-lymphocyte
population (or a subpopulation). The timing of this event could be orchestrated by, for
example, administration of the appropriate factor.
Currently, treatment with AZT
must be prolonged because the triggering of latent HIV (DNA) to destroy itself is mainly
dependent on G0/G1 switches generated by random
antigenic signals. At one point in time only one cell (or a small group of cells), becomes
permissive for HIV replication. The cell, with its integrated HIV DNA, is then destroyed.
AZT prevents the liberated viruses replicating and establishing latency in fresh host
cells. However, if all host T-lymphocyte were activated synchronously by an appropriate
concentration of an appropriate growth factor, all the integrated HIV DNA molecules would be
destroyed with their host cells. Furthermore, all
liberated RNA viruses could be prevented from replicating in previously uninfected cells
by a short and intensive concomitant course of AZT. For this purpose, the effectiveness of
AZT might be increased by combining AZT with drugs (e.g. 5-fluorodeoxyuridine; 25, 31),
which deplete intracellular pools of natural AZT competitors and reduce feedback
inhibition of the enzyme which is rate-limiting for incorporation of AZT (32).
While much
remains to be learned about the normal signals affecting cell population size homeostasis,
there are indications that this 'one shot' approach to AZT
therapy is feasible. Matsuyama et al (33) have suggested that a suitable polyclonal 'growth factor' might be tumor necrosis factor-alpha
(TNF-alpha).
This can activate host cell NFkB-like factors which, in turn, can activate latent HIV
(34, 35). Treatment with TNF-alpha alone would be expected to accelerate the progression of
AIDS (36). Concomitant treatment with AZT might turn
this progression to the advantage of the host. |
HIV
infection of non-lymphoid tissues
The above discussion is concerned with eliminating the AIDS virus
from the T-lymphocyte population and thus preventing or correcting
acquired immune deficiency. The virus can also persist in various other tissues, thus
constituting a reservoir from which reinfection of T-lymphocytes might occur
(16).
Cotherapy with AZT and an appropriate end-cell specific cytokine or growth factor, to
convert the cells to a state permissive for viral growth, might eliminate this reservoir
(37, 38). |
Acknowledgements. I thank Dr. P Ford for helpful comments
on the manuscript. These studies were supported by grants from the American
Foundation for AIDS Research (AMFAR) and Queen's
University.
References
1. Fischl MA, Richman DD, Grieco
MH, Gottlieb MS, Volherding PA, Laskin OL, Leedom JM, Groopman JE, Mildvan D, Schooley RT,
Jackson GG, Durack DT, Phil D, King D. Aids Collaborative Working Group. The efficiency of
AZT in the treatment of patients with AIDS and AIDS-related complex. New Eng J Med 317, 185, 1987.
2. Yarchoan R, Mitsuya H, Myers CE, Broder S. Clinical pharmacology of
3'-azido-2', 3'-dideoxythyniidine and related dideoxynucleosides. New Eng J Med 321, 726, 1989.
3. Yarchoan R, Broder S. Development of antiretroviral therapy for AIDS
and related disorders. New Eng J Med 316,
557, 1987.
4. Mitsuya H. Broder S. Strategies for antiviral therapy in AIDS. Nature 325, 773, 1987.
5. Peterlin BM, Luciw PA. Replication of the human
immunodeficiency virus: strategies for inhibition. Biotechnol
6, 794, 1988.
6. Haseltine WA. Development of antiviral strategies for the treatment
of AIDS: strategies and prospects. JAIDS 2,
311, 1989.
7. Forsdyke DR. Further implications of a theory of immunity. J Theor. Biol 52, 187, 1975.
8. Schwartz RH. Acquisition of immunologic self-tolerance. Cell 57, 1073, 1989.
9. Bluestone JA, Cron RQ, Cotterman M, Houlden BA, Matis LA. Structure
and specificity of T cell receptor gamma-delta on MHC antigen-specific CD3+, CD4-, CD8- T
lymphocytes. J Exp Med 168, 1899, 1988.
10. Porcelli S. Brennar M B, Greenstein J L, Balk S P, Terhorst C.
Bleicher PA. Recognition of cluster of differentiation antigens by human CD4, CD8,
cytolytic T-lymphocytes. Nature 341, 447,
1989.
11. Vidovic D. Rogfic M, McKune K, Guerder S, Mackay C, Dembic Z. Qa-1
restricted recognition of foreign antigens by a gamma-delta T-cell hybridoma. Nature 340, 646, 1989.
12. Wallis V J. Leuchards E, Chauduri H, Davies JS. Studies in
hyperlymphoid mice. Immunology 38 38, 163,
1979.
13. Rocha B, Dautigny N, Pereira P. Peripheral T-lymphocytes: expansion
potential and homeostatic regulation of pool sizes and CD4/CD8 ratios in vivo. Eur J Immunol. 19, 905, 1989.
14. Miller RA, Sturman 0. T-cell repopulation from functionally
restricted splenic progenitors. J. Immunol 133,
2925, 1984.
15. Fauci AS. The human immunodeficiency virus: infectivity and
mechanism of pathogenesis. Science 259, 617, 1988.
16. Levy JA. Mysteries of HIV: challenges for therapy and prevention. Nature 333, 519, 1988.
17. Bloom BR, Senik A, Stoner G, Ju M, Nowakowski M, Kano S, Jiniinez
L. Studies on the interactions between viruses and lymphocytes. Cold Spring Harb Conf Quant Biol. 41, 73, 1976.
18. McDougal JS, Mawle A, Cort SP, Nicholson JK, Cross GD,
Scheppler-Campbell JA, Hicks D, Sligh J. Cellular tropism of the human retrovirus
HTLVI/LAV. Role of T-cell activation and expression of the T4 antigen. J Immunol 135 135, 3151, 1985.
19. Nabel G. Baltimore D. An inducible transcription factor activates
expression of HIV in T-cells. Nature 326,
711, 1987.
20. Forsdyke DR. cDNA cloning of mRNAs which increase rapidly in human
lymphocytes cultured with concanavalin-A and cycloheximide. Biochem
Biophys Res Com 129, 619, 1985.
21. Greene WC, Bohnlein E, Ballard W. Immunol
Today 10, 272, 1989.
22. Rosenberg ZF, Fauci AS. AIDS Res Hum
Retrovir 5, 1, 1989.
23. Cullen B, Green WC. Regulatory pathways governing HIV-1
replication. Cell 58, 423, 1989.
24. Lim D, Maas WK. Reverse transcriptase in bacteria. Molec. Microbiol 3, 1141, 1989.
25. Forsdyke DR, Scott FW. Evidence for non-convergence of de novo and
salvage pathways of purine deoxyribonucleotide synthesis, p. 177, in Cell Compartmentation and Metabolic Channeling. Ed.
(L Nover, F Lynen, K Moher) Elsevier Pub Co, Amsterdam, 1980.
26. Mathews CK, Moen LK, Wang Y, Sargent RG. Intracellular organization
of DNA precursor biosynthetic enzymes. TIBS 139,
394, 1988.
27. Preston BD, Poiesz BF, Loeb LL. Fidelity of HIV-1 reverse
transcriptase. Science 242, 1168, 1988.
28. Loeb LA. Endogenous Carcinogenesis: Molecular Oncology into the
Twenty-first Century. Cancer Res. 49, 5489,
1989.
29. Temin HM. Retrovirus variation and evolution. Genome 31, 17, 1989.
30. Furman PA, Fyfe JA, St Clair MH, Weinhold K, Rideout JL, Freeman
GA, Lehrman SN, Bolognesi DP, Broder S, Mitsuya H, Barry DW. Phosphorylation of
3'-azido-3'deoxythymidine and selective interaction of the 5'-triphosphate with HIV
reverse transcriptase. Proc Natl Acad Sci USA 83,
8333, 1986.
31. Sjostrom DA, Forsdyke DR. Isotope-dilution analysis of
rate-limiting steps and pools affecting the incorporation of thymidine and deoxycytidine
into cultured thymus cells. Biochem J 138,
253, 1974.
32. Forsdyke DR. Treatment of pneumocystic pneumonia in AIDS. New Eng J Med 318, 989, 1988.
33. Matsuyama T. Hamamoto Y, Soma G, Mizuno D, Yamamoto N, Kobayashi N.
Cytocidal effect of tumor necrosis factor on cell s chronically infected with HIV:
enhancement of HIV replication. J Virol 63,
2504, 1989.
34. Lowenthal JW, Ballard DW, Bohnlein E, Greene WC. Tumor necrosis
factor alpha induces proteins that bind specifically to kappa- B-like enhancer elements
and regulate interleukin-2 receptor alpha chain expression in primary human T-lymphocytes.
Proc Natl Acad Sci. USA 86, 2331, 1989.
35. Osbome L, Kunkel S, Nabel GJ. Tumor necrosis factor alpha and
interieukin-I stimulate the HIV enhancer by activation of nuclear factor kappa-B. Proc Natl Acad Sci USA 86, 2336, 1989.
36. Folks TM, Clouse KA, Justement J, Rabson A, Duh E. Kehri JH, Fauci
AS. Tumor necrosis factor alpha induces expression of HIV in a chronically infected T-cell
clone. Proc Natl Acad Sci USA 86, 2365,
1989.
37. Folks TM, Justement J, Kinter A, Dinarello CA, Fauci AS, Cytokine-induced
expression of HIV-I in a chronically infected promonocyte cell line. Science
238, 800, 1987.
38. Perno CF, Yarchoan R, Cooney DA, Hartman NR, Webb DS, Hao Z, Mitsuya H, Johns DG,
Broder S. Replication of HIV in monocytes. J Exp Med
169, 933, 1989.
Some quotations
from Tae-Wook Chun et al. (1998)
"The presence of this latent reservoir of HIV is of considerable concern
since these cells remain as a potential source of
reactivation of viral replication."
"Since it is likely that ...
infected cells die upon activation of virus and that HAART prevents spread of virus to
adjacent cells, the observation that this combination of cytokines [IL6, TNF-alpha, IL2]
can markedly induce viral replication in this reservoir may have important implications
for the activation-mediated diminution of the latent
reservoir of HIV in patients receiving HAART."
"Since cytokines alone can
reactive HIV-1 replication in latently infected, resting, CD4+ cells, and since these
cells probably die upon reactivation, it is conceivable that a
strategy of administration of cytokines together with HAART might result in a diminution
of this reservoir of latently infected cells".
"Given the relatively long
half-life of these latently infected, resting CD4+ T cells,
and the fact that they are constantly within an environment capable of providing the
stimuli for reactivation, it is not unreasonable to explore
strategies aimed at deliberately diminishing the size of this pool of cells.
In this regard, since cytokine-mediated induction of HIV-1 replication in these cells with
subsequent release of virus probably results in death of the cell, and since the presence
of HAART in vitro prevents spread of released virus, it is conceivable that in vivo administration of cytokines together with HAART may have
such an effect."
|
Some quotations from
David Ho (1998)
"Infectious HIV-1 persists
latently in resting, memory CD4 lymphocytes in a post-integrated form despite 1 to 2 years
of combination therapy [i.e. AZT or similar compounds + protease inhibitors]. This latent
reservoir of HIV-1, denoted "L" , ... represents the major documented hurdle to virus eradication,
although other ... viral sanctuaries may exist."
"5 to 7 years of continuous,
completely inhibitory therapy will be necessary to eliminate L. Treatment
interruptions that permit HIV-1 replication to resume will rapidly restore the size of L.
For larger pool sizes ... more than 10 years of continuous treatment will be required. A
treatment duration this protracted is unacceptable
because of the complexity, toxicity and cost of the current drug regimes... ."
"Infectious proviruses are
undoubtedly harbored within a diverse population of resting CD4 lymphocytes with memory
for a large array of exogenous antigens. Thus, administration
of a limited set of antigens is unlikely to activate a sufficient number of these cells to
replicate and thereby facilitate their rapid death. On the other hand, the
use of a large panel of antigens would be impractical.
What about the administration of cytokines?
Interleukin-2 (IL-2) alone is not expected to activate resting [G0] lymphocytes because
such cells do not express the appropriate high affinity receptor. However, mixtures of
certain cytokines, such as IL-2, IL-6 and tumor necrosis
factor, have been shown to activate resting T cells in vitro. "
|
Steve Bunk's article from The Scientist
7 Dec 1998
New Weapon Attacks Latent HIV
Reservoirs. (Click Here)
| The Scientist
14, no.2 page 6, 24th January 2000 (Click Here): COMMENTARY (with
copyright permission from the publisher Alexander Grimwade)
HIV: A Grouse-shooting Analogy
By Donald R. Forsdyke
The Hot Papers
article1 of Dec. 6 on the failure of various combinations of antibiotics to
eradicate latent HIV gives the false impression that AIDS researchers were not aware of
this possibility. ("Scientists are still grappling with the questions raised by this
sobering discovery.")
Doctors learn at medical school the fundamental rule that antibiotics should be given for
short periods in
adequate doses to
destroy all pathogens and prevent the emergence of resistant strains. As soon as it was
appreciated that AIDS was caused by a retrovirus, it was predictable that antibiotics
alone would be unlikely to work. Retroviruses usually have a latency option and are highly
prone to mutate.
Thus, future therapy would have to be rather like grouse-shooting; one would need guns to
shoot the birds and beaters to flush them out. To rid an area of grouse neither alone
suffices. The combination is lethal.2
Accordingly, the research agenda for AIDS had to develop along two lines:
- (1) research on viral growth and
replication to find antibiotics such as AZT, which would hit the virus
"on the wing," and
- (2) research on latency to
find drugs such as TNF-alpha, which would flush out viral reservoirs.
A short and inexpensive period of treatment
should then suffice (expense being a particularly important factor in Third World
countries).
Unfortunately, our research systems do not operate this way.3,4 Antibiotics
were emphasized rather than latency-disrupting drugs. When AZT did not
come up to expectations and resistant strains emerged, the call came for bigger and better
guns, rather than for beaters. If regular guns won't work, add the howitzers! If that
combination does not work, add the mortars! And to forestall criticism, call it
"highly active anti- retroviral therapy" (HAART)!
The quite predictable
consequence is that we have a variety of initially highly effective
antibiotics to which HIV is now resistant. Thankfully, HAART can increase
the life span of patients, but these same patients, by virtue of the resistant strains
they harbor, remain as potentially dangerous sources of infection. When latency-disrupting
drugs eventually emerge (and let us hope the work on IL-2 is not just hype), it may be too
late. HIV may be totally resistant.5,6
Part of the problem is a refusal to use the word "antibiotic"
in the context of HIV treatment. Instead, patients are subject to
antiretroviral therapy with "agents" or "drugs." Furthermore, it is argued that "therapy for HIV-1 disease can be
viewed in a way that is similar to treatment for cancer," instead of similar to
treatment of other pathogen-caused diseases.7
References
1. S. Bunk with comments by D.D. Richman and R.F. Siliciano, Hot Papers, The Scientist,
13[24]:22, Dec. 6, 1999.
2. D.R. Forsdyke, "Programmed activation of T lymphocytes: a theoretical
basis for short term treatment of AIDS with Azidothymidine," Medical
Hypotheses, 34: 247, 1991.
3. D.R. Forsdyke, "A systems analyst asks about AIDS research
funding," Lancet, 2:13824,
1989.
4. D.R. Forsdyke, "Bicameral grant review: how a systems analyst with AIDS
would reform research funding," Accountability in
Research, 2:23741, 1993.
5. D.R. Forsdyke, www.queensu.ca/academia/forsdyke/aids.htm
6. D.R. Forsdyke, Tomorrow's Cures Today?
Newark, Harwood Academic, 2000. (Click Here)
7. R.J. Pomerantz, "Residual HIV-1 disease in the era of highly active
antiretroviral therapy." New England Journal of
Medicine, 340:16724, 1999. |
Comment on the above Commentary:
2000-02-02 email
"Regarding your article in The Scientist:
I think that the 'reason for refusal to use the word
"antibiotic" in
the context of HIV treatment' is that many virologists think that
term is incorrect. Many microbiologists, and most virologists I
know, consider an "antibiotic" to refer strictly to
an antibacterial
agent (never mind that its etymological meaning can be broader).
For agents against viruses, "antiviral" is the
standard term (and
against fungi, antimycotic, etc.).
[Prefers to remain anonymous]
Professor
[
]
Department of Genetics
University of [ ]"
USA
A similar point was made in
The
Scientist March 20th (Click Here),
which was briefly replied to in the April issue: (Click Here)
Reply to Comment:
Definition of "Antibiotic"
Thank you for your comment concerning "the standard
term". Webster's Dictionary
defines "antibiotic", when used as an adjective, as something "tending to prevent, inhibit, or destroy life." As you imply, a
strict etymological interpretation would regard lions as antibiotic with respect to
humans, and Popeye as antibiotic with respect to spinach. Clearly the word has to be
understood in context.
In the present discussion this context relates to chemicals which
are antibiotic with respect to microorganisms which can invade the body of a host
organism, such as man. Antibiotics are not "antiseptics"
or "disinfectants" which can sterilize at the
surface or outside of a host, but are usually not tolerated within host tissues.
Thus, in the present context I would define the noun "antibiotic"
as a chemical [of natural or
synthetic origin] which, [usually at low
concentrations], inhibits microorganisms of some type within a host
organism, while not unacceptably interfering with the life of that organism.
[This does not exclude the possibility that the chemical will also inhibit the
microorganisms outside the host (e.g. on a Petri dish) but, by virtue of being
tolerated within the body of the host, the chemical is an antibiotic not an antiseptic.
Most modern antibiotics work at low concentrations, and are toxic to the host at
high concentrations. It is possible that in future we might find a chemical which works
only at high concentrations and these concentrations are not toxic to the host. So
concentration should not be in the definition.]
Since, historically, many of the early antibiotics worked only against
bacteria ("antibacterial antibiotics"), it is
easily to fall into the trap of thinking that antivirals ("antiviral
antibiotics") are different. If we think of them as in a separate,
non-antibiotic, category, then the lessons
we have learned about antibiotic usage (i.e. mode of use to prevent resistant strains
emerging) can easily get disregarded (the point of my Commentary).
Unfortunately, the Webster's
Dictionary (1976) definition has archaic aspects when
defining the noun as "a substance produced by a microorganism and able in dilute solution to
inhibit or kill another microorganism". It gets it right when referring to the
target of an antibiotic as "another microorganism"
(virus, bacterium, fungus, protozoan, etc.), but is wrong in postulating that antibiotics
are necessarily "produced by a microorganism".
Yes, historically, many antibiotics were isolated from
microorganisms (e.g. penicillin produced by a fungal mould), but also many
were synthesized by the chemist (the arsenicals for the bacterium causing syphilis,
and the sulphonamides, which were effective against a variety of bacteria). The
list of synthetic antibiotics now includes AZT (antiviral antibiotic) and modified forms of penicillin
(antibacterial antibiotics). Antibiotic forms initially
derived by purification from microorganisms are now
being chemically synthesized and further modified.
This history can be reversed. Among the
antibiotics now available only
through chemical synthesis, in future we may find some which are synthesized by some
organism, perhaps an organism yet to be discovered. Including the source of antibiotics,
and/or the type of microorganisms they attack, in the definition, is archaic. Our
nomenclature must move with the times in order not to confuse ourselves, our students, and
our patients.
Donald Forsdyke
Another gospel: American
Heritage Dictionary 1996.
"NOUN:
A substance, such as penicillin or streptomycin, produced by or derived from certain
fungi, bacteria, and other organisms, that can destroy or inhibit the growth of other
microorganisms. Antibiotics are widely used in the prevention and treatment of infectious
diseases.
ADJECTIVE:
1. Of or relating to antibiotics. 2. Of or relating to antibiosis. 3. Destroying life or
preventing the inception or continuance of life". For
more on this matter, see: Bentley, R.&
Bennett, J. W. (2003) What is an antibiotic?
revisited. Advances in
Applied Microbiology 52, 303-331. |
The Truth Continues to Dawn
[commentary
circa 2000]
"However, the most worrisome reservoir consists of latently infected resting
memory CD4+ T cells carrying integrated HIV-1 DNA. Definitive demonstration of the
presence of this form of latency required development of methods for isolating extremely
pure populations of resting CD4+ T cells and for demonstrating that a small fraction of
these cells contain integrated HIV-1 DNA that is competent for replication if the cells
undergo antigen-driven activation.
Most of the latent virus in resting CD4+ T cells is
found in cells of the memory phenotype. The half-life of this latent reservoir is
extremely long (44 months). At this rate, eradication of this reservoir would require over
60 years of treatment. Thus, latently infected resting CD4+ T cells provide a mechanism
for life-long persistence of replication-competent forms of HIV-1, rendering unrealistic
hopes of virus eradication with current antiretroviral regimens.
The extraordinary
stability of the reservoir may reflect gradual reseeding by a very low level of ongoing
viral replication and/or mechanisms that contribute to the intrinsic stability of the
memory T cell compartment. Given the substantial long-term toxicities of current
combination therapy regimens, novel
approaches  to eradicating this latent reservoir are urgently
needed." to eradicating this latent reservoir are urgently
needed."
T. Pierson, J. McArthur & R. F. Siliciano. (2000) Annual
Reviews of Immunology 18, 665-708. |
And Dawn...
| In October 2000 it was eventually admitted
that: "In
cases with apparent complete HIV suppression by HAART, viral rebound after cessation of
therapy could have originated from the activation of virus from the latent reservoir."
(Zhang et al. 2000).
In a major concession to the viewpoint
advanced on this web-page it was stated:
"Several
research goals assume paramount importance.
First, it is
critical to determine whether additional viral reservoirs exist.
Second, it is
important to understand the nature and source of the ongoing virus production that is seen
in most patients on HAART.
Finally, novel
approaches are needed to eliminate latently infected cells which clearly represent a very
serious barrier to HIV-1 eradication."
That such research goals
"assume" importance only
by the year 2000 is of note. The readers of these pages
may infer that individuals such as the authors cited above were among those rejecting
grant applications from researchers who, a decade or more before, had considered it quite
obvious that these goals were of paramount importance.
Their
goals did not get support, not because of scientific logic, but because of the mind-set of
the well-intentioned people who held the political high ground, but all-too-often
could not see beyond their noses.
There is nothing new in this, as the example of
diphtheria immunization in the early decades of the 20th century has shown (Forsdyke,
2000). Amazingly, in
2003 Dr. Siliciano declared that: "It
is difficult to envision any targeting mechanism that will allow specific elimination of
this reservoir."(Click Here). Of course, he is not
required to "envision." He just has to read the scientific
literature (see above)!
The implications of this go far beyond the management of natural diseases.
Currently, the greatest threat to humankind seems to be overt or terrorist
warfare conducted not with nuclear weapons, but with biological weapons. A
nation which uses the peer-review process, as it currently (2001)
operates, to select those who give it advice on biomedical matters, may
not fare well in confrontation with a nation which has adapted the
peer-review process to identify those (e.g. Irvine Page, Szent-Gyorgyi,
Erwin Chargaff) who can see beyond their noses.
Forsdyke, D. R. (2000) Tomorrow's Cures Today? How to
Reform the Health Research System. Harwood Academic, Newark.
Siliciano, J. D. & Siliciano, R. F. (2000) Latency and viral persistence in HIV-1
infection. J. Clin. Invest. 106,
823-824.
Zhang, L., Chung, C., Hu, B-S., He, T., Guo, Y., Kim, A. J., Skulsky, E., Jin, X.,
Hurley, A., Ramratnam, B., Markowitz, M. & Ho, D. D. (2000) J.
Clin. Invest. 106, 839-845. |
Bioinformatic Analysis of Retroviruses (Click Here).
Deals with the origin
of retroviral species (with implications for speciation in general), and notes a
region of high conservation just upstream of GAG, which is vital for
dimerization and packaging and is thus a potential target
sites for therapeutic agents (e.g. antisense oligonucleotides).
Codon Choice in
Retroviruses (Click Here)
There are 20
amino acids and 61 codons, and one amino acid can have more than one codon. All genes in a
particular genome tend to use the same set of codons. In his "genome hypothesis" Grantham concluded that
differences in choice of codons by different species reveal the presence of fundamental
genomic forces. We ignore these at our peril.
Death
While Seeking the Origin of AIDS (Click Here)
William
D. Hamilton, the biologist whose work was popularized in Dawkins' The
Selfish Gene, went to the jungle to find out for himself.
More of the Same in South Africa
(Click Here) By putting all
their "eggs" into the immunization "basket" the Medical Research Council of South Africa
became part of the
problem not the solution to the problem.
The Drug Companies Win the "Image Game" and Promote the Spread
of AIDS (Click Here)
|
