Honoured President, my lords and gentlemen, -- It is to me the very greatest honour that I have been summoned here by your most highly esteemed Society, which for more than two centuries has represented and still represents the centre of the scientific life of England, in order that I may deliver the Croonian Lecture. I consider I am not so much personally concerned in the honour that you bestow on me, and that I shall not err if I see in it a recognition of the scientific path which I, in company with many others, have sought to follow, and which in your eyes suffices to place the field in which I work on a footing alongside of exact science. It is an extreme pleasure for me to have the privilege of addressing so many medical colleagues with whom for so many years I have been bound in close ties of friendship, and who have always been the first to welcome and to give recognition to the results of my work. |
Since Jenner made his great discovery of the protective action of vaccinia against small-pox, a century has passed away. During these years that terrible scourge of mankind has been almost completely eradicated from the civilised world. |
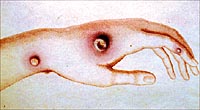 |
The beneficial consequences of Jenner's discovery are so evident to all who have any wish to properly appreciate them, that one wonders why, during so great a portion of the long period of 100 years, they were allowed to stand alone, without any endeavour being made to induce an artificial immunity in the case of other infectious diseases. This is all the more remarkable because Jenner's discovery demonstrated in their entirety those essential principles which, in later times, have been established for other infectious diseases. In the first place, it was shown that by the use of an attenuated virus, which of itself was non-injurious to the organism, it was possible to ward off the disease caused by the virulent virus. Jenner also established -- what is most important from the practical point of view -- that by the inoculation of the weakened poison there was produced not only an immediate, but also an enduring, protection. |
That Jenner's discovery remained so isolated was due essentially to the fact that the theoretical conceptions of the cause and nature of infectious diseases made no advance during the subsequent decades; indeed, it would be an interesting topic for some historian of medicine to trace, step by step, the gradual advance in the knowledge of infectious diseases during the past century.[For Rinderpest 1866 (Click Here)] Schwann's classical investigations must be regarded as the first link in the long chain. Schwann it was who, in an unusually brilliant manner, first demonstrated that the decomposition of organic bodies in the processes of fermentation and putrefaction was never spontaneous, but constantly arose through the agency of micro-organisms coming from without. This line of investigation reached its zenith in the fundamental work of Pasteur, of which the first and the greatest result -- Lister's method of wound treatment -- worked a revolution in surgery. Then followed the profound investigations of Koch on Anthrax, and the pure cultivation of the most important pathogenic bacteria. |
The work of Pasteur and of Koch afforded the first basis on which the study of artificial immunity could be again undertaken. The possibility of voluntarily producing a number of the most important infectious diseases of men and animals, and of modifying at will pure cultivations of bacteria, either, according to Jenner's precedent, by passage through the animal body, or otherwise in artificial culture media, laid the foundation on which advancement could proceed. Pasteur himself was the first, after Jenner, to produce an artificial immunity by using an attenuated virus; and he was also able to introduce the procedure to some extent into practice with most beneficial results. Still, the theoretical explanations of all these facts lagged far behind their practical effects. The very able investigations of Metchnikoff and his theory of phagocytosis were, to many investigators, inconclusive. |
Immune bodies (Antibodies, Antikorper)
With Behring's discovery, that in the blood serum of animals immunised against diphtheria and tetanus, there were contained bodies which were able to specifically protect other animals against the toxines of these diseases, an altogether new factor was introduced into the question. This remarkable discovery seemed at one stroke to open up an entirely new and extremely promising prospect of immunising mankind against the majority of the infectious diseases. It was, therefore, somewhat disappointing when there did not follow, on the successful practical application of diphtheria antitoxic serum, a rapid succession of similar achievements. It may with truth be said, that during recent years there has been somewhat of a standstill in the following-out of a work at first so enthusiastically received. By purely empirical methods, e.g., by the production and use of sera of very great antitoxic value, the results attained showed no improvement. Better success was only to be hoped for when, by an accurate knowledge of the theoretical considerations underlying the question of immunity, explanations of the previous ill-success were forthcoming. Impelled by these considerations I laboured for years trying to shed some light into the darkness that shrouded the subject. |
In all exact work with chemical bodies -- for only as such can we regard the toxines produced by the living bacteria -- the first desideratum in the investigation is the exact numerical determination of action and counteraction. The words of the gifted natural philosopher Clerk Maxwell, who said that if he were required to symbolize the learning of our time he would choose a metre measure, a clock and a kilogramme weight, are equally apposite in reference to progress in the field of inquiry in which we are at present interested. And so, at the very beginning of my theoretical work on immunity, I made it my first task to introduce measures and figures into investigations regarding the relations existing between toxine and antitoxine. From the outset it was clear that the difficulties to come were extremely great. The toxines, i.e., the poisonous products of bacteria, are unknown in a pure condition. So great is their potency, that we are obliged to assume that the strongest solid (feste) poisons which are obtained by precipitating toxic bouillon with ammonium sulphate, represent nothing more than indifferent materials, peptones and the like, to which the specific toxine attaches itself in mere traces beyond the reach of weighing; for up to the present time, by the purely chemical methods of weighing and measuring, it has impossible to ascertain anything as to their presence or the intensity of their action. |
Their presence [the presence of toxins] is only betrayed by the proof of their specific toxicity on the organism. For the exact determination e.g., of the amount of toxine contained in a culture fluid, the essential condition was that the research animals used should exhibit the requisite uniformity in the susceptibility to the poison. Uniformity is not to be observed in the reaction of the animal body to all toxines. Fortunately in the case of one important body of this nature, viz., the diphtheria toxine, the conditions are such that the guinea-pig affords for investigations the degree of accuracy necessary in purely chemical work. For other toxines this accuracy in measuring the toxicity cannot be attained. It was necessary for me to try to eliminate, as far as possible, the varying factor of the animal body, and bring the investigations more nearly into line with the conditions necessary for experiments of a chemical nature. In the course of these endeavours it was shown that it was possible to obtain in a comparatively simple manner an insight into the theoretical considerations necessary to a proper understanding of immunity, by means of test-tube experiments with suspended animal tissues. The relations were simplest in the case of red blood corpuscles. On them, outside the body, the action of many blood poisons, and of their antitoxines, can be most accurately studied, e.g., the actions of ricin, eel-serum, snake-poison, tetanus toxine, etc. In an experiment of this kind, in which are employed a series of test-tubes containing definite quantities of suspended blood corpuscles, each test-tube represents as it were a research animal, uniform in any one series, and one that can be reproduced at will. By means of these test-tube experiments, particularly in the case of ricin, I was able, in the first place, to determine that they yielded an exact quantitative representation of the course of the processes in the living body. The demonstration of this fact formed the basis of a more extended application of experiments of this nature. It was shown that the action of toxine [antigen] and antitoxine [antibody] took place quantitatively as in the animal body. Further, these experiments yielded a striking series of facts of importance for the theoretical valuation of the reaction between toxine and antitoxine. It was proved in the case of certain toxines -- notably tetanus toxine -- that the action of antitoxines is accentuated or diminished under the influence of the same factors which bring about similar modifications in chemical processes -- warmth accelerates, cold retards the reaction, and this proceeds more rapidly in concentrated than in dilute solutions. These facts, first ascertained by means of test-tube experiments, have since been confirmed by Behring and Knorr for tetanus within the animal body, and by Martin and Cherry in the case of snake-venom.* {Footnote: Note during revision.-- The credit of first drawing attention to these points belongs to Professor Fraser, who, as far back as 1896, carried out extraordinarily precise experiments on the conditions of neutralisation in respect both of time and of amount of snake-poison and anti-venin (Lecture, Royal Institution, March 20, 1896).} The knowledge thus gained led easily to the inference that to render toxine innocuous by means of antitoxine was a purely chemical process, in which biological processes had no share. Yet again insurmountable obstacles seemed to present themselves to this conclusion. |
Discrepancy between assays in vitro (in test tubes) and in vivo (in the body)
It must be postulated that in chemical processes the bodies sharing in the action react with one another in definite equivalent quantities. This proposition appeared, however, not to hold in the case of the action of antitoxins on diphtheria toxine. When, in the case of Diphtheria toxines of different stocks, that quantity of toxine bouillon which is exactly neutralised by a certain definite quantity of diphtheria antitoxine (the official German immunity unit, as laid down for the control examination of sera) was determined, so that every trace of toxic action was abolished, the figures obtained were not in accord. Of one toxine bouillon 0.2 c.c., of another 2.5 c.c., were so neutralised by one immunity unit. Such a relation need not have given rise to surprise, because it was well known that the diphtheria bacillus, according to outside circumstances, yields in the bouillon very different quantities of toxine. It was therefore allowable to infer that the different quantities of toxine bouillon, which were saturated by one immunity unit, were exact expressions of the toxicities of the various bouillons, or, to use other words, indifferently whether the bouillon was strongly or feebly toxic, the same multiple of the minimal lethal dose would be constantly neutralised by one immunity unit, so that in every case the law of equivalent proportions would hold good. But when looked into more closely, the relations showed themselves to be by no means so simple. In what manner could one obtain a satisfactory estimation of the strength of a toxine? As the constant factor in such an estimation, it was only possible to proceed from a previously determined standard reaction in the case of a definite species of animal, and so we came to regard as the "toxic unit" that quantity of toxic bouillon which exactly sufficed to kill, in the course of four days, a guinea-pig of 250 grammes weight. When we employed this standard unit, or "simple lethal dose," to estimate the amount of toxic bouillon neutralised by one "immunity unit," the facts which presented themselves were far more surprising than it was possible to have foreseen at the outset. These results were, that of one toxine, perhaps 20, of a second, perhaps 50, and of yet a third, it might be 130 simple lethal doses were saturated by one immunity unit. Since, however, we had previously assumed that the simple lethal dose alone afforded a standard on which reliance could be placed in determining the combining relations of toxine and antitoxine, it appeared from these results that the neutralisation of toxines by antitoxines did not follow the law of equivalent proportions, and, notwithstanding all earlier work in agreement with such a conception of the action, we were obliged to conclude that between toxine and antitoxine a purely chemical affinity did not exist. The seemingly inexplicable contradiction between the results just stated and previous work was very soon explained. When the neutralisation point of toxine and antitoxine was investigated for one and the same sample of poison, the following results were obtained. Immediately on its preparation, fresh from the incubator, it was found that one immunity unit neutralised a c.c. of toxic bouillon, and this quantity represented beta simple lethal doses. When the same toxic bouillon was examined after a considerable interval, the remarkable fact was discovered that exactly alpha c.c. of the toxic bouillon were again neutralised by one immunity unit; but that these alpha c. c. now represented only beta minus x simple lethal doses. It therefore followed that the toxic bouillon had retained exactly the same combining affinity [in vitro], but possessed feebler toxicity[in vivo]. From this it was evident that the toxic action on animals and the combining capacity with antitoxine represented two different functions of the toxine, and that the former of these had become weakened, while the latter had remained constant. |
Two chemical groups on toxine, haptophore and toxiphore
Treated from the chemical standpoint, this circumstance was most simply explained by assuming that the toxine was characterised by the possession of two different combining groups: one, which may be designated haptophore, conditions the union with antitoxine, while the other group, which may be designated toxophore, is the cause of the toxic action. From the constancy of the combining capacity, and the diminution in the toxicity, it was to be inferred that:
If we now designated a toxine molecule, of which the toxophore group is destroyed, but its haptophore group retained, as "toxoid," then the above-described process will represent the quantitative progress of the conversion of the toxine molecules into toxoid molecules. Such a toxoid molecule has the same quantitative combining affinity for antitoxins as the original toxine molecule, in spite of the disappearance of toxicity to the animal body. In other words, the affinity of the haptophore group for the antitoxine is absolutely independent of the existence of a toxophore group. Also, in the original toxine molecule, both groups must be to such a degree non-related or independent of one another, that a mutual reaction between them does not take place. This conception of the constitution of diphtheria toxine, after more extensive, very exact, and much varied experimentation, based on its partial neutralisation by antitoxine, has been confirmed in the completest manner possible. At this time it would be superfluous for me to enter into all the details pertaining to these investigations. It need only be remarked that in principle the same relations have been established for tetanolysine by Madsen, for snake-poison by Meyers, and for the milk-curdling ferment by Morgenroth. |
Distribution of chemicals in the body?
The separation of the characteristic atom groups of the toxine molecule into a haptophore and a toxophore group, afforded not merely a satisfactory chemical explanation of the process of neutralisation: the possession of the knowledge of the existence of these groups yielded us, at the same time, the key to the nature of the toxic property of toxines, and to the mystery of the origin of the antitoxines themselves. After it had been established by the already described method, that the toxine molecule was possessed of a definite haptophore group, which accounted for its capacity to enter into combination with other bodies, it was immediately necessary to inquire into the question whether, and if so to what degree, this group entered into the causation of the symptoms of illness. That chemical substances are only able to exercise an action on the tissue elements with which they are able to establish an intimate chemical relationship is a conception of a general nature, which has been entertained since the birth of scientific medicine. It is astonishing, almost astounding, that this axiom, of which the theoretical importance has been so long recognised, and which has served indeed as the first ground for certain therapeutical procedures, should as a matter of fact have played in the building up and furtherance of scientific pharmacology a role so insignificant in proportion to its great importance. In glancing through the modern text-books of pharmacology, with rare exceptions, as, e.g., Stokvis, one finds absolutely no mention, of the distribution of drugs in the organism, a matter which is of so much moment for arriving at a true comprehension of the relations existing between pharmacological action, location in the organism, and chemical constitution. As a matter of fact, the methods for obtaining any knowledge of the exact distribution of drugs in the body are as yet very imperfect. Even if we can prove that certain alkaloids are again recognisable as being, e.g., present in the brain, we are but little further advanced in our knowledge of the process, because we cannot determine in which cells and which system of fibres the alkaloid is localised. I may say, indeed, that as yet the investigation of the laws pertaining to the minute distribution of a chemical substance in the body is only possible when, as in the case of coloured bodies, these are at once recognisable by the eye. But that it is possible at once to draw conclusions of therapeutic importance from the laws governing the distribution was shown in the case of methylene-blue, in which I was able, knowing its distribution in the body, to anticipate for it certain antineuralgic and antimalarial properties which were both established by subsequent investigation. It may be permitted me to call to mind that in malaria methylene blue is especially of service in the case of persons who, on account of susceptibility, cannot be treated with quinine, and that in the hands of Koch it has shown itself of eminent value in haemoglobinuric fever, since as opposed to quinine it exercises no destructive action on the erythrocytes. If we are not able to discover the principles governing the localisation of common chemical bodies, which can be used in suitable quantities in chemical purity, and which chemical and other reactions render perceptible, it was a priori very unlikely that efforts directed to locating the toxines, which are potent in the slightest traces, and which are bodies we have no means of rendering perceptible to our senses, would be anything else than absolutely without result. |
Coinjection of toxin with antitoxin needed for protection
But that this is not so, has been shown by experiments carried out by Professor Donitz, in the Steglitz Institute, to which, on account of their great importance, I shall refer somewhat extensively. When a rabbit receives a suitable dose of diphtheria or tetanus toxine injected directly into the circulation, the animal remains for many hours well, and then begins to show symptoms of illness, which gradually increase till they end in death. In order to arrive at an explanation of the incubation period, Donitz determined the amount of antitoxine which, injected intravenously immediately after the toxine, absolutely neutralised the latter. This neutralising dose is able to render all the toxine circulating in the blood innocuous. When, however, the neutralising dose so determined was injected not immediately, but seven or eight minutes after the injection of the toxine, death occurred from tetanus exactly as if no antitoxine had been given. Part of the toxine, equal at least to the minimal lethal dose, must within this time have disappeared from the blood, in which it would have been neutralised, and passed over to the tissues, especially to the brain. The experiments of Donitz were afterwards confirmed by an investigation conducted in quite a different manner by Heymans, who showed that a research animal from which the blood had been removed immediately after the injection of the minimal lethal dose of tetanus toxine, and replaced by transfusion of fresh blood, succumbed from typical tetanus. In this case, therefore, in that brief interval of time the minimal lethal dose of toxine had passed through the walls of the vessels and been taken up by the tissues. Regarding the nature of the processes here concerned, a satisfactory explanation was also afforded by the experiments of Donitz. It admitted of demonstration that the toxine held in the tissues could still be withdrawn from them, if not the simple neutralising dose were injected, but larger quantities of the same. The quantity necessary was greater in proportion as the interval elapsing after the injection of the toxine was longer. However, after a definite period was exceeded, all possible doses of antitoxins, even the very greatest, were impotent, notwithstanding that the animal at the time of the injection of the antitoxine had not developed any symptoms. Since a very great number of other chemical substances, narcotics, alkaloids, and other neurotropic bodies, were not in a position to withdraw the toxine once deposited in the central nervous system, and as the property to do so was solely the characteristic of the specific antitoxins, one was obliged to come to the conclusion that the union between the toxine and the tissues, which could only be overcome by means of a specific chemically-related antagonising agent, must itself depend on a chemical combination. One was therefore forced to accept the idea that the central nervous system, that is to say certain ganglion cells in it, possessed atom groups resembling those of the antitoxins, in having a maximum affinity for tetanus poison. The predilection of the nervous system for tetanus toxine, the rapid union of the toxine with the nervous tissue, the gradual onset of the symptoms and their long duration could only be explained by the existence of such toxophile groups. The statement of Donitz that the tetanophile atom groups are in the guinea-pig essentially confined to the central nervous system, whereas in the case of other species, especially rabbits, these are also present in other organs, is one of prominent importance. The beautiful experiments of Roux on intracerebral injection of toxine have yielded absolute confirmation of the statement of Donitz. Roux found in guinea-pigs that the same dose of tetanus toxine was lethal, whether given by intracerebral or by subcutaneous injection; for rabbits, however, the lethal dose was twenty times greater subcutaneously than it was in intracerebral injection. This can only be explained in the terms of Donitz's observation, viz., that in the case of direct injection of the toxine into the brain, the toxophile atom groups there present at once seize on all the toxine, while when the toxine is administered through the blood stream, the toxophile groups present in other organs also take up the toxine in equivalent quantities. In the case of rabbits the absorption of the toxine in this way is very considerable: indeed of twenty parts only one part finds its way into union with the nervous system.
One may therefore rightly assume that these toxophile protoplasmic groups in reality serve normal functions in the animal organism, and that they only incidentally and by pure chance possess the capacity to anchor themselves to this or that toxine. [The following article in this web-page shows how, 50 years later, this conceptual problem was overcome by Talmage and Burnet.] |
Antigens (toxines) are large and metabolizable
The first thought suggested by this assumption [that toxophile groups serve a normal function] was that the atom groups referred to must be concerned in tissue change; and it may be well here to sketch roughly the laws of cell metabolism. Here we must in the first place draw a clear line of distinction between those substances which are able to enter into the composition of the protoplasm, and so are really assimilated, and those which have no such capacity. To the first class belong a portion of the food-stuffs par excellence; to the second almost all our pharmacological agents, alkaloids, antipyretics, antiseptics, &c. How is it possible to determine whether any given substance will be assimilated in the body or not ? There can be no doubt that assimilation is in a special sense a synthetic process -- that is to say, the molecule of the food-stuff concerned enters into combination with the protoplasm by a process of condensation involving loss of a portion of its water. To take the example of sugar, in the union with protoplasm, not sugar itself as such but a portion of it comes into play, the sugar losing in the union some part of its characteristic combining reactions. The sugar behaves here as it does, e.g., in the glucosides, from which it can only be obtained through the agency of actual chemical cleavage. The glucoside itself yields no trace of sugar when extracted in indifferent solvents. In a quite analogous manner, the sugar entering into the constitution of albuminous bodies (glycoproteids) cannot be obtained by any method of extraction; at least, not until chemical decomposition has previously taken place. It is therefore generally easy, by means of extraction experiments, to decide whether any given combination in which cells take part is, or is not, a synthetic one. If alkaloids, aromatic amines, antipyretics, or aniline dyes be introduced into the animal body it is a very easy matter, by means of water, alcohol, or acetone, according to the nature of the body, to remove all these substances quickly and easily from the tissues. This is most simply and convincingly demonstrated in the case of the aniline dyes. The nervous system stained with methylene blue, or the granules of cells stained with neutral red, at once yield up the dye in the presence of alcohol. We are therefore obliged to conclude that none of the foreign bodies just mentioned enter synthetically into the cell complex; but are merely contained in the cells in their free state. The combinations into which they enter with the cells, and notably with the not really living parts of them (Kupffer's paraplastic portions), are very unstable, and correspond usually only to the conditions obtaining in solid solutions, while in other cases only a feeble salt-like formation takes place. I myself in 1887 placed on a sure footing the fact that the nervous system and the fatty tissues allow of alkaloids and aniline dyes being mechanically shaken out of them, as in the poison-detection process of Stas and Otto. |
Pharmacological agents are generally non-antigenic
Hence with regard to the pharmacologically active bodies in general, it was not allowable to assume that they possessed definite atom groups, which entered into combination with corresponding groups of the protoplasm. This corresponds, as I may remark beforehand, with the incapacity of all these substances to produce antitoxines in the animal body. We must therefore conclude, that only certain substances, food-stuffs par excellence, , are, endowed with properties admitting of their being, in the previously defined sense, chemically bound by the cells of the organism. We may regard the cell quite apart from its familiar morphological aspects, and contemplate its constitution from the purely chemical standpoint. We are obliged to adopt the view, that the protoplasm is equipped with certain atomic groups, whose function especially consists in fixing to themselves certain food-stuffs, of importance to the cell-life. Adopting the nomenclature of organic chemistry, these groups may be designated side-chains. |
We may assume that the protoplasm consists of a special executive centre (Leistungs-centrum) in connection with which are nutritive side-chains, which possess a certain degree of independence, and which may differ from one another according to the requirements of the different cells. And as these side-chains have the office of attaching to themselves certain food-stuffs, we must also assume an atom-grouping in these food-stuffs themselves, every group uniting with a corresponding combining group of a side-chain. The relationship of the corresponding groups, i.e., those of the food-stuff, and those of the cell, must be specific. They must be adapted to one another, as, e.g., male and female screw (Pasteur), or as lock and key (E. Fischer). From this point of view, we must contemplate the relation of the toxine to the cell. We have already shown that the toxines possess for the antitoxines an attaching haptophore group, which accords entirely in its nature with the conditions we have ascribed to the relation existing between the food-stuffs and the cell side-chains. And the relation between toxine and cell ceases to be shrouded in mystery if we adopt the view that the haptophore groups of the toxines are molecular groups fitted to unite, not only with the antitoxines, but also with the side-chains of the cells, and that it is by their agency that the toxine becomes anchored to the cell |
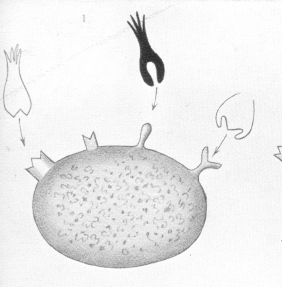 |
We do not, however, require to suppose that the side-chains, which fit with the haptophore groups of the toxines, i.e., the side-chains which are toxophile, represent something having no function in normal cell economy. On the contrary, there is sufficient evidence that the toxophile side-chains are the same as those which have to do with the taking up of the food-stuffs by the protoplasm. |
The toxines are, in opposition to other poisons, of highly complex structure, standing in their origin and chemical constitution in very close relationship to the proteids and their nearest derivatives. It is, therefore, not surprising if they possess a haptophore group corresponding to that of a food-stuff. Alongside of the binding haptophore group, which conditions their union to the protoplasm, the toxines are possessed of a second group, which, in regard to the cell, is not only useless but actually injurious. And we remember that in the case of the diphtheria toxine there was reason to believe that there existed alongside of the haptophore group another and absolutely independent toxophore group. Now for certain cellular elements of the body it can be proved in the test-tube that between these tissues and certain toxines an "anchoring" process takes place exactly similar to that between toxine and antitoxins. Wassermann first demonstrated this in the case of the brain substance. In a mixture of tetanus toxine and broken-down fresh guinea-pig brain the latter so bound or "anchored" the toxine, that not only was the surrounding fluid toxine-free, but the brain substance laden with the tetanus toxine had also lost its own toxic action, and so the mixture when injected into an animal was borne without any harm. The deduction is, that in this case, a chemical union between the brain substance and the tetanus toxine had taken place, and this was of so firm a nature that on introduction into the body the union was not broken up and therefore the toxine remained innocuous. The brain of the normal animal had, in keeping with my theory, acted exactly like a real antitoxine. There are present in the brain, i.e., in the gang lion cells, tetanophile protoplasmic groups, which unite themselves with the toxine. The presence of such groups is the necessary preliminary and cause of the poisonous action of the tetanus toxine in the living animal. That the process here was not one of simple absorption is proved by the fact that, if the group concerned was destroyed by heat, the brain substance became as incapable of removing the toxine as an emulsion of any other organ of the guinea-pig. |
Properties of haptophore and toxophore groups
As has been said, the possession of a toxophile group by the cell is the necessary preliminary and cause of the poisonous action of the toxine. This can be most sharply demonstrated in the case of certain blood poisons, viz., the haemolysines, which exercise a solvent action only on such red blood corpuscles as are able to unite chemically with them. The union with the red corpuscles can be proved, and one has here the great advantage of dealing with living and intact red blood cells instead of broken-down cellular material. Under these conditions it is easy to determine the quantitative relations of the union. If we now regard the action of the toxines with which we are concerned in accordance with the views we have just been discussing, we are obliged to conclude that these are only in a position to act prejudicially on the organism if they are able, by means of their haptophore groups, to anchor themselves to the side-chains of the cells of organs essential to life. If the cells of these organs lack side-chains fitted to unite with them, the toxophore group cannot become fixed to the cell, which therefore suffers no injury, i.e., the organism is naturally immune. One of the most important forms of natural immunity is based upon the circumstance, that in certain animals the organs essential to life are lacking in those haptophore groups which seize upon definite toxines. If, for example, the ptomaine occurring in sausages, which for man, monkeys, and rabbits is toxic in excessively minute doses, is for the dog harmless in quite large quantities, this is because, the binding haptophore groups being wanting, the ptomaine cannot, in the dog, enter into direct relation with organs essential to life. We see, then, that the haptophore groups act especially in bringing definite areas of the cell within the sphere of influence of the toxophore group. In the behaviour of the haptophore and toxophore groups there exists a difference essentially great, as we have already pointed out when referring to the work of Donitz and Heymans. The haptophore group exercises its activity immediately after injection into the organism, while in all toxines -- with the, perhaps, solitary exception of snake-venom -- the toxophore group comes into activity after the lapse of a longer or shorter incubation period, which may, e.g., in the case of diphtheria toxine, extend to several weeks. It is in the highest degree interesting that it is possible, by voluntarily influencing certain of the outside conditions, to exclude absolutely the action of the toxophore group. Courmont has shown that frogs, when kept at a temperature lower than 20o C., manifest no sign of tetanus, even after very large doses of tetanus toxine, but they succumb to fatal tetanus if they are placed in surroundings of a higher temperature. Dr. Morgenroth, working in my Institute, has throw light on this behaviour by proving that in frogs maintained in cold surroundings the tetanus toxine is fixed in their central nervous system, and that the absence of action at lower temperatures can only be explained by the toxophore group of tetanus toxine having its action restricted within a certain temperature minimum, while independent of this the haptophore group exercises its action on the nervous system at all temperatures. |
Origin of antibodies (Antikorpor)
The theory above developed allows of an easy and natural explanation of the origin of antitoxines [see images 1-6]. In keeping with what has already been said, the first stage in the toxic action must be regarded as being the union of the toxine by means of its haptophore group to certain ''side-chains" of the cell protoplasm. This union is, as animal experiments with a great number of toxines show, a firm and enduring one. |
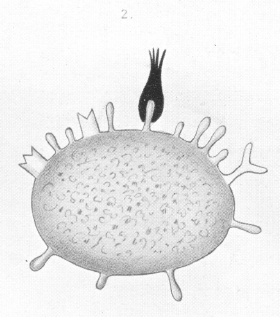 |
The side-chain involved, so long as the union lasts, cannot exercise its normal physiological nutritive function -- the taking up of definite foodstuffs. It is as it were shut out from participating, in the physiological sense, in the life of the cell. We are therefore now concerned with a defect which, according to the principles so ably worked out by Professor Carl Weigert, is repaired by regeneration. These principles, in fact, constitute the leading conception in my theory. |
|
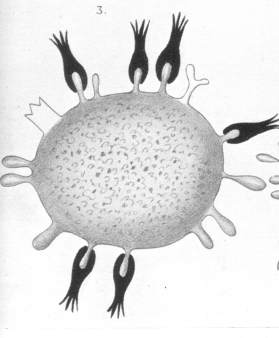 |
If, after union has taken place, new quantities of toxine are administered at suitable intervals and in suitable quantities, the side-chains, which have been reproduced by the regenerative process, are taken up anew into union with the toxine, and so again the process of regeneration gives rise to the formation of fresh side-chains. In the course of the progress of typical systematic immunisation, as this is practised in the case of diphtheria and tetanus toxine especially, the cells become, so to say, educated or trained to reproduce the necessary side-chains in ever-increasing quantity. |
As Weigert has confirmed by many examples, this, however, does not take place as a simple replacement of the defect; the compensation proceeds far beyond the necessary limit; indeed, over-compensation is the rule. Thus the lasting and ever-increasing regeneration must finally reach a stage at which such an excess of side-chains is produced that, to use a trivial expression, the side-chains are present in too great a quantity for the cell to carry, and are, after the manner of a secretion, handed over as needless ballast to the blood. |
|
Regarded in accordance with this conception, the antitoxines represent nothing more than side-chains reproduced in excess during regeneration, and therefore pushed off from the protoplasm, and so coming to exist in a free state. . With this explanation the phenomena of antitoxins formation lose all their strange, one might say miraculous, characters. |
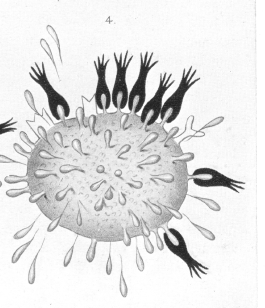 |
I have deemed it advisable to represent by means of some purely arbitrary diagrams (Plates 6 and 7 [images from which are reproduced here]) the views I have expressed regarding the relations of the cell considered in the manner I have been describing. Needless to say, these diagrams must be regarded quite apart from all morphological considerations, and as being merely a pictorial method of presenting my views on cellular metabolism, and the method of toxine action and antitoxine formation during the process of immunisation. In the first place our theory affords an explanation of the specific nature of the antitoxines, that tetanus antitoxine is only caused to be produced by tetanus toxine, and diphtheria antitoxine through diphtheria toxine. This very specific nature of the affinity between toxine and cell is the necessary preliminary and cause of the toxicity itself. Further, our theory makes it easy to understand the long-lasting character of the immunity produced by one or several administrations of toxine, and also the fact that the organism reacts to relatively small quantities of toxine by the production of very much greater quantities of antitoxine. |
|
By the act of immunisation, certain cells of the organism become converted into cells "secreting" antitoxine at the same rate as this is excreted. New quantities of antitoxine are constantly produced, and so throughout a long period the antitoxine content of the serum remains nearly constant. The secretary nature of the formation of antitoxines has been very strikingly illustrated by the beautiful experiments of Salmonson and Madsen, who have shown that pilocarpine, which augments the secretion of most glands, also occasions in immunised animals a rapid increase in the antitoxine content of the serum. |
The production of antitoxines must, in keeping with our theory, be regarded as a function of the haptophore group of the toxine, and it is therefore easy to understand why, out of the great number of alkaloids, none are in a position to cause the production of antitoxines. Conversely, indeed, I recognise in this incapacity of the alkaloids, in opposition to the toxines, to produce antitoxines, a further and salient proof of the truth of the deduction I have previously based on chemical grounds, that the alkaloids possess no haptophore group which anchors them to the cells of organs. To formulate a general statement, the capacity of a body to cause the production of antitoxine stands in inseparable connection with the presence of a haptophore atomic group. In the formation of antitoxine the toxophore group of toxine molecule is, on the contrary, of absolutely no moment. But the toxoid modifications of the toxines, in which the haptophore group of the toxine is retained, while its toxophore group has ceased to be active, possess the property of producing antitoxines. Indeed, in some cases of extremely susceptible animals immunity can only be attained by means of the toxoids, and not by the too strongly acting toxines. The toxoids are certainly able to cause the production of antitoxines. To quote an example, it is hardly possible in an animal, which, like the guinea-pig, has all the tetanophile groups confined to the cells of the central nervous system, to produce immunity by means of the unaltered tetanus toxine, whereas this is attained with extraordinary rapidity and ease by means of its toxoids. The symptoms of illness due to the action of the toxophore group, therefore, play no part in the production of antitoxine. On the contrary, we may consider that the severe symptoms, which indicate injury to the cell-life, disturb the regenerative functions, and thus hinder or entirely frustrate the course of the immunisation process. I have from the first adopted this view, and it was simply a misunderstanding when Knorr, who has been all too soon taken from the field of his labours, affirmed that, according to my theory, sickness of the cell constituted the necessary condition precedent to the new formation and pushing-off of side-chains. |
If I am not altogether deceived, the toxoids, where there is a question of producing active immunization (and this will always be the case when the immunization concerns human beings), are destined to play an important role in practical medicine. |
The world of antigens other than toxines
In their theoretical relations the toxoids are also of far-reaching interest, in that they provide a transition to that immunisation which can be called forth by substances which would a priori be considered entirely devoid of toxic character, and which are sometimes, like the autochthonous ferments (i.e., those normally present in blood), products of normal cell-life, and in some cases food-stuffs proper. |
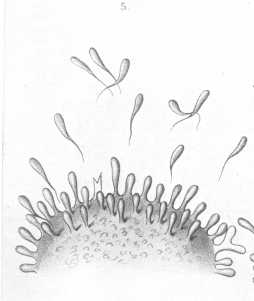 |
Thus, Dr. Morgenroth, working in my laboratory, has proved that the rennet ferment, if introduced in great quantities into the organism, behaves exactly like a real toxine, in that it causes the production of a typical anti-rennet, which, up to a certain limit, accumulates in proportionally greater quantity, the greater the injected doses. Here, however, we have to do with processes which are altogether within the region of the normal, as is most clearly shown in certain animals, e.g., the horse, in the blood serum of which there is normally present a quantity of anti-rennet, equal to that attained in the goat only after a systematic immunisation carried on for months. The rennet ferment present naturally in the body of the horse is the cause of this great formation of anti-rennet. According to Bordet's experiments, if injections of milk be given to animals, their serum acquires thereby the capacity to cause flocculent curdling. This action is seemingly rigidly specific because (according to Morgenroth's experiments) the body produced by the injection of goat's milk, coagulated goat's milk, but not human or cow's milk. The behaviour is also similar when different kinds of albumin, e.g. the sera of different animals or the white of egg, are injected. There appear constantly in the serum of the animal so treated new substances -- specific coagulines -- which act only in a specific manner, i.e., precipitate only the form of albumin injected. Thus there are produced, by the injection of common food-stuffs, typical "Antikorper," which unite with the substances used to occasion their production, and form with them insoluble combinations. |
|
My investigations have shown me that in the blood of animals which have not been subjected to any treatment we must accept the presence of a number of normal bodies analogous to the "Antikorper," having their origin in the most widely diverse organs, and representing nothing more than nutritive side-chains, which in the course of the normal nutritive processes have been developed in excess and pushed off into the blood. From all these considerations I think myself warranted in concluding that the formation of antitoxines lacks all the characters of that purposeful, intelligently directed, and remarkable process which it at first seemed to be, and that it is to be regarded merely as a process analogous to those constituting an essential portion of the normal metabolism of the organism. We must admit that the majority of the foodstuffs and of the intermediate products of tissue-change must be able to cause the production and throwing-off of nutritive side-chains. It may be that the new formation only takes place to a limited extent, and that the replacement of any side-chains which have been shut out from their physiological function is all that is accomplished; but the formation may occur in greater proportions, may become excessive, and therefore lead to the presence of "Antikorper " in the blood. |
|
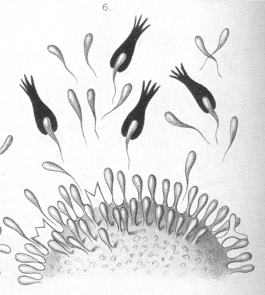 |
In this way is easily explained the fact of the occurrence in the normal blood serum of antitoxines and of bodies inimical to bacteria, without the animals having ever been brought into relation with the corresponding toxines or bacteria. Here I need only refer to the fact that diphtheria antitoxine is not uncommonly present in normal horses and in men who have never suffered from diphtheria. |
Particularly weighty in this connection are the observations that have been made on horses, because, on the one hand, these animals never suffer from diphtheria, and, on the other hand, Cobbett has brought forward experimental proof that this normally occurring antitoxine corresponds absolutely as to its properties with the antitoxins produced by artificially immunising. The conclusion, therefore, is that in the body of the normal horse certain substances may be present which possess side-chain affinities similar to those of the diphtheria toxine, and which, therefore, are quite as capable as the latter are of taking possession of the cell side-chains, and occasioning the regeneration and pushing-off of these from the cells; in other words, of causing the presence of an actual diphtheria antitoxine in a normal animal. Such occurrences direct attention to the possibility of producing immunity in some cases by the administration of definite food-stuffs. Perhaps we have in some such peculiarity of feeding and tissue-change the explanation of the fact so difficult to understand, viz., that individuals of the same race and species react in such diverse manners to the same infection. Certainly we are very far remove from the solution of this important question, which, as yet, has scarcely assumed a tangible form. Still it is our duty to strive with tenacity to overcome the difficulties which surround this point, bearing in mind the words of your illustrious countryman, Francis Bacon: "Sunt certe ignavi regionum exploratores, qui, ubi nil nisi coelum et pontus videtur, terras ultra esse prorsus negant." |
|
Opposition to side-chain theory
I have now laid before you the fundamental facts which up to the present constitute our knowledge in the field pertaining to immunity, and which can be most easily and successfully explained through the agency of "the side-chain theory." I wish in a few words to dispel some erroneous ideas which have been advanced in opposition to this theory. Roux has shown that very small quantities of tetanus toxine, if injected directly into the brain, cause the death of the animal. Roux assumes that such an occurrence is not compatible with my theory. Roux is of opinion that according to my theory the brain must be quite immune against tetanus toxine, as the toxophile side-chains of the brain-cells must be identical with the antitoxine, and therefore exercise an immediate protective action. Experiment showing quite the reverse, the theory is overthrown. Roux came to this incorrect conception through an erroneous conception of antitoxine. The toxophile side-chains of the brain cells draw directly to themselves the toxine molecules, and, according to my theory, are thus a necessary preliminary condition of the illness. The toxophile groups are therefore really inducers of the action of the poison, and not its preventives. Those toxophile groups which, like the antitoxines present in the serum, are able to lay hold of toxine immediately on its entry into the blood, and so to divert it from organs essential to life, can alone be regarded as being possessed of any antitoxic action in the true sense of the word. I may be allowed to call to mind Weigert's excellent simile of iron and the lightning conductor. Iron attracts electricity and is therefore used as a lightning conductor. Great masses of iron present in buildings give rise to, or increase, the risk of their being struck by lightning, and the metal only becomes protective against lightning when it is so employed that the electricity is conducted away outside the building. It would never occur to anyone to speak of great masses of iron machinery present in buildings as if they were lightning conductors. It is equally unreasonable to speak of the antitoxic property of the brain cortex, in which the toxophile groups are present in great quantity, but also retain their relations with the nerve-cells. When this really considerable misunderstanding is eliminated from Roux's results these become entirely confirmatory of my views, and it is difficult to understand how, subsequent to Weigert having placed the matter in so clear a light, the beautiful experiments of Roux can be utilised by another eminent authority as a means of combating my theory. |
Immunization with whole microorganisms generates many antibody species
Much more complex than in the cases hitherto discussed are the conditions when, instead of the relatively simple metabolic products of microbes, the living micro-organisms themselves come to be considered, as in immunisation against cholera, typhoid, anthrax, swine fever, and many other infectious diseases. There then come into existence alongside of the antitoxines, produced as a result of the action of the toxines, manifold other reaction products. This is because the bacterium is a highly complicated living cell, of which the solution in the organism yields a great number of bodies of different nature, in consequence of which a multitude of "Antikorper" are called into existence. Thus we see, as a result of the injection of bacterial cultures, that there arise alongside of the specific bacteriolysines, which dissolve the bacteria, other products, as, for example, "coagulines" (Kraus, Bordet), i.e., substances which are able to cause the precipitation of certain albuminous bodies contained in the culture fluid injected; also the so-much discussed "agglutinins" (Durham, Gruber, Pfeiffer), the antiferments (von Dungern), and no doubt many other bodies which we have not yet recognised. It is by no means unlikely that each of these reaction products finds its origin in special cells of the body; on the other hand, it is quite likely that the formation of any single one of these bodies is not of itself sufficient to confer immunity. Thus in case of the introduction of bacteria into the body we have to do with a many-sided production of different forms of "Antikorper," each of which is directed only against one definite quality or metabolic product of the bacterial cell. Accordingly, in recent times, the practice of using for the production of immunisation definite toxic bodies isolated from the bacterial cells has been more and more given up, and for this purpose it is now regarded as important to employ the bacterial cells as intact as possible. The beautiful results obtained for plague by Haffkine, and quite recently by Wright in your own country for typhoid fever, have been arrived at in this way. |
Specificity of bacteriolysis and haemolysis
The most interesting and important substances arising during such an immunising process are without doubt the bacteriolysines, in the investigation of which Pfeiffer has done such yeoman's service. How really wonderful it is that after the introduction of the cholera vibrio into the animal body a substance is formed endowed with the power of dissolving the cholera vibrio, and that vibrio only! This seemingly purposeful and novel phenomenon seems at first sight to have nothing to do with those forces which are normally at the disposal of the organism. It was of the greatest importance to explain the origin of these substances from the standpoint of cellular physiology. The solution offered very considerable difficulties, and was first attained when, instead of bacteriolysines, haemolysines came to be employed in experiments. Haemolysines are peculiar toxic bodies, which destroy red blood corpuscles by dissolving them. Haemolysines may occur in a normal blood when they exercise a solvent action on the red blood corpuscles of other species, or they may be artificially produced, in which case, after an animal has undergone a process of immunisation against the blood corpuscles of another species, there appear in the serum haemolysines which destroy the kind of blood corpuscles employed in the production of the immunity. In their essential characters they are absolutely comparable with the bacteriolysines: but they possess over them the great advantage that they admit of being employed in test-tube experiments, and thus afford opportunity for exact quantitative work altogether independent of the variability of the animal body. Belfanti and Carbone first discovered the remarkable fact that horses which have been treated with the blood corpuscles of rabbits contain in their serum constituents which are poisonous for the rabbit, and for the rabbit only. While the serum of the normal horse, to the quantity of 60 c.c., could be intravenously injected without harm to the rabbit, a very few c.c. of serum from horses previously so treated with rabbit's blood, proved fatal. |
Bordet showed shortly thereafter, that in the case quoted there was present in the serum a specific haemolysine which dissolved the blood corpuscles of the rabbit. He also proved that these haemolysines -- as had already been shown by Buchner and Daremberg in the case of similarly acting bodies which are present in normal blood -- lost their solvent property on being maintained during half an hour at a temperature of 55oC. Bordet added, further, the new fact, that the blood-solvent property of these sera which had been deprived of solvent power by heat, the solvent action could be restored if certain normal sera were added to them. By this important observation an exact analogy was established with the facts of bacteriolysis as elicited by the work of Pfeiffer, Metchnikoff, and Bordet. In the work on the Pfeiffer phenomenon of bacteriolysis, it had already been ascertained that the solution of bacteria by specific bacteriolysines was brought about by the combined action of two different bodies: one which was specific, arose during the immunisation and was stable; and another, a very unstable body, which was present in normal serum. In collaboration with Dr. Morgenroth, I have sought in regard to this question, for which haemolysis offered prospects favourable to experimentation, to make clear the mechanism concerned in the action of these two components -- the stable, which maybe designated "immune body," and the unstable, which may be designated "complement" -- which, acting together, effect the solution of the red blood corpuscles. For this purpose, in the first place, solutions containing either only the "immune body" or only the "complement" were brought in contact with suitable blood corpuscles, and after separation of the fluid and the corpuscles by centrifugalising, we investigated whether these substances had been taken up by the red blood corpuscles or remained behind in the fluid. The proof of its location in the one position or in the other was readily forthcoming, since to restore to the haemolysine its former activity, it was only necessary to add to the "immune body" a fresh supply of "complement," or to the "complement" a fresh supply of "immune body," in order that the presence of the haemolysine in its integrity might be shown by the occurrence of solution of the blood-cells. The experiments proved that, after centrifugalising, the "immune body" is quantitatively bound to the red blood corpuscles, and that the "complement," on the contrary, remains entirely behind in the fluid. The presence of the two components in contact with blood corpuscles only occasions the solution of these at higher temperatures, and not at 0oC. And an active haemolytic serum (with "immune body" and "complement" both present) having been placed in contact with red blood corpuscles and maintained for a while at 0oC., it was found after centrifugalising that, under these circumstances also, the "immune body" had united with the red blood corpuscles, but that the "complement" remained in the serum. This experiment showed that both components must, at a temperature of 0oC., have existed alongside of one another in a free condition. But when analogous experiments were undertaken at a higher temperature it was found that both components were retained in the sediment. |
|
These facts can only be explained by making certain assumptions regarding the constitution of the two components, i.e., of the "immune body" and the "complement." In the first place, two haptophore groups must be ascribed to the "immune body," one having a great affinity for a corresponding haptophore group of the red blood corpuscles, and with which at lower temperatures it quickly unites, and another haptophore group of a lesser chemical affinity, which at a higher temperature becomes united with the "complement" present in the serum. |
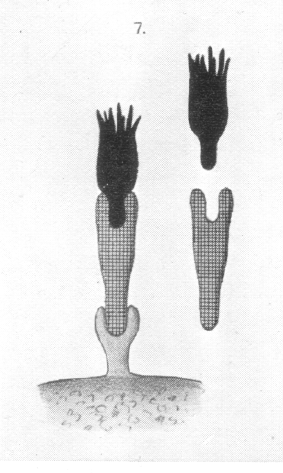 |
Therefore, at the higher temperature, the red blood corpuscles will draw to themselves those molecules of the "immune body" which in the fluid have previously become united with the "complement." In this case the "immune body" represents in a measure the connecting chain which binds the complement to the red blood corpuscles, and so brings them under its deleterious influence. Since under the influence of the "complement" -- at least, in the case of the bacteria, -- appearances are to be observed (for example, in the Pfeiffer phenomenon) which must be regarded as analogous to digestion, we shall not seriously err if we ascribe to this "complement" a ferment-like [enzymic] character. It is obvious that when the normal serum of one animal possess haemolytic action on the blood of another, the component of the haemolysine which here unites with the red blood corpuscle and forms the connecting link between it and the "complement" which is essential to the occurrence of solution, cannot, in the absence of any preceding process of immunisation, be designated "immune body." In its characteristics and action, however, it only differs from this in occurring naturally, and may well be designated "intermediate body" (Zwischenkorper; ["natural antibody"]). It may here be stated that the constitution of a haemolysine is graphically represented in fig. 7, Plate 7 [see above]. Very important for the conclusion that only with the assistance of the "intermediate body" [natural antibody] or of the "immune body" [induced antibody] can the "complement," which leads to the solution, become united with the blood corpuscle, is the following experiment. The serum of the dog has very considerable solvent action upon guinea-pig's blood, but loses this property if warmed. If dog's serum, thus rendered inactive by warming, is brought into contact with suspended corpuscles of guinea-pig's blood, these are not dissolved; but, if to such a mixture there be also added guinea-pig serum, i.e., the serum normal to these red blood corpuscles, the erythrocytes are at once dissolved. Here the only explanation is that the [natural antibody] "intermediate body," which possesses a specific affinity for guinea-pig erythrocytes, and is present in the inactive dog's serum, is able to seize on one of the many "complements" present in guinea pig's serum, with the result that the "complement" which cannot normally attach itself to the corpuscles, comes now to, exercise its destructive influence. We see at the same time from this experiment that the haemolysines occurring naturally, obey the same laws as those produced through the process of immunising. In fact, for them also, in a great number of instances, precisely similar behaviour has been demonstrated. The character of the specific union made it possible to find solutions for a number of important questions. In the first place, regarding the multiplicity of the haemolysines, which occur normally in serum, it is well known that numerous sera are able to dissolve blood corpuscles of different species. For example, serum of the dog dissolves blood corpuscles of the rabbit, guinea-pig, rat, goat, sheep, &c. The complex nature of these haemolysines has been already indicated. |
|
Antibody specificity shown by adsorption to cells
Another question arises whether, in a serum that is capable of such manifold action, there is present one single haemolysine that destroys different red blood cells, or whether a whole series of haemolysines come into action, of which one is adapted to guinea-pig blood, another to rabbit blood, etc. The solution of this question may be approached in another way. The serum may be rendered inactive by heat, and then placed in contact with red blood corpuscles of a given kind. Then, supposing, for example, that rabbit blood has been employed, it is found that if the fluid is freed from the erythrocytes by centrifugalisation and the "complement" afterwards added, it is no longer in a position to dissolve rabbit blood, but has not suffered any impairment of its action on other kinds. By this method of elective absorption it is proved that the normally occurring haemolysines which chain the blood corpuscles of the rabbit to themselves [cause them to agglutinate], are specifically adapted to this purpose. If with suitable adjustment of conditions similar experiments be conducted with other kinds of blood, results are obtained which force us to the conviction that in such a serum acting on various kinds of blood there are present absolutely different [natural antibodies] "intermediate bodies" (analogues of the "immune bodies"), of which each one is specific for one kind of blood, i.e., one is adapted for rabbit's blood, a second for calf's blood, etc. Dr. Morgenroth and I have in some cases, indeed, succeeded in proving that the "complements" which are adapted to fit themselves to these [natural antibodies] "intermediate bodies", and occur in normal sera, differ among themselves. If we reflect that in normal blood, in addition to these different haemolysines, there are besides a long series of analogous bodies, agglutinines of very different kinds, bacteriolysines, enzymes, anti-enzymes, we are brought more and more to the conviction that the blood serum is the carrier of substances innumerable as yet little known or conceived of.. |
Having obtained a precise conception of the method of action of the lysines of the serum -- of the haemolysines, and thereby also of the bacteriolysines -- it becomes possible for us to attempt to solve the mystery of the origin of these bodies. I have in the beginning of this lecture fully developed the "side-chain theory," according to which the antitoxines [antibodies] are merely certain of the protoplasm, "side-chains," which have been produced in excess and pushed off into the blood. |
|
 |
The toxines [antigens], as secretion products of cells, are in all likelihood still relatively uncomplicated bodies; at least by comparison with the primary and complex albumins of which the living cell is composed. If a cell of the organism has, with the assistance of an appropriate "side chain," fixed to itself a giant molecule, as the proteid molecule really is, then, with the fixation of this molecule, there is provided one of the conditions essential for the cell nourishment. |
| Such giant molecules cannot at first be utilised by the cells, and are only made available when, by means of a ferment-like [enzymic] process, they are split into smaller fragments. This will be very effectually attained if, figuratively speaking, the "tentacle" or grappling arm of the protoplasm possesses a second haptophore group adapted to take to itself ferment-like material out of the blood fluid. Through such complex organisation, by which the "tentacle" acts also as the bearer of a ferment-functioning group, this group is brought into close relation with the prey destined to be digested and assimilated. For such appropriate arrangements, in which the "tentacular" apparatus also exercises a digestive function -- if it be permissible to pass from the abstract to the concrete -- we find analogies in the different forms of insectivorous plants. Thus it has been known since the famous researches of Darwin that the tentacles of Drosera secrete a proteid-digesting fluid. If, we now recognise that the different lysines only arise through absorption of highly complex cell material -- such as red blood corpuscles or bacteria -- then the explanation, in accordance with what I have said, is that there are present in the organism "side-chains" of a special nature, so constituted that they are endowed not only with an atomic group by virtue of the affinities of which they are enabled to pick up material, but also with a second atomic group, which, being ferment-loving in its nature, brings about the digestion of the material taken up. Should the pushing-off of these "side chains" be forced, as it were, by immunisation, then the "side-chains" thus set free must possess both groups, and will therefore in their characteristics entirely correspond to what we have placed beyond doubt as regards the "immune-body" of the haemolysine. In this manner is simply and naturally explained the astonishingly specialised arrangement that, through the introduction of a definite bacterium into the body, something is produced which is endowed with the power of destroying by solution the bacterium which was administered and no other. This contrivance of the organism is to be regarded as nothing more than a repetition of a process of normal cell-life, and the outcome of primitive wisdom on the part of the protoplasm. |
|
In conclusion, I wish hastily to touch on only a few points. First, to direct attention to the fact that the immunising sera produced by the administration of bacteria are sometimes limited in their operation to certain animal species, and are much more inconstant in their action than are the antitoxines. Sobernheim, in the laboratory of C. Fraenkel, found that the anthrax serum obtained by immunising German marmots (Hamster) protected this species, even in small doses; but was absolutely without action for rabbits. Kitt had a precisely similar experience with symptomatic anthrax. This circumstance is easy to understand, if the complex nature of the lysines be borne in mind. The lysine, be it bacteriolysine or haemolysine (i.e., "immune body" + "complement"), possesses altogether three haptophore groups, of which two belong to the "immune-body" and one to the "complement." Each one of these haptophore groups can be bound by an appropriate "anti-group". Three anti-groups are thus conceivable, any one of which, by uniting with one of the haptophore groups of the lysine, can frustrate the action of the lysine. To my mind, of these three possible "Antikorper," that one which can lay hold of the haptophore group of the "complement," and so prevent this from uniting with the "immune body," is the most important. Dr. Morgenroth and I have experimentally succeeded in producing such bodies by processes of immunisation, and in proving that they unite with the "complement" (anticomplement). Dr. Neisser at the Steglitz Institute sought to find an explanation of Sobernbeim's experiments. He was able to determine that anthrax serum failed in mice, even if great quantities of fresh sheep's serum (i.e., containing excess of "complement") were at the same time introduced. The failure in this case appears to be due, on the one hand, to the destruction, in the body of the mouse, of the "complement" present in the sheep's serum, and, on the other hand, to the fact that the "immune body" yielded by the sheep does not find in mouse serum an appropriate new "complement." From this it appears, that in the therapeutic application of antibacterial sera to man, therapeutical success is only to be attained if we use either a bacteriolysine with a "complement" which is stable in man ("anthropostabile complement"), or at least a bacteriolysine, the "immune body" of which finds in human serum an appropriate "complement." The latter condition will be the more readily fulfilled the nearer the species employed in the immunisation process is to man. Perhaps the non-success which as yet has attended the employment of typhoid and cholera serum will be converted into the contrary if the serum be derived from apes and not taken from species so distantly removed from man as the horse, goat, or dog. However this may be, the question of the provision of the appropriate "complement" will come more and more into the foreground, for it really represents the centre round which the practical advancement of bacterial immunity must turn. |
Antibodies against mammalian cells
A second and at present much-discussed question, is the immunising of the organism against elements standing biologically much higher in the scale than erythrocytes and much less foreign to the body than those exceedingly lowly organisms, the bacteria. I refer here to the production of "Antikorper" against cells of the higher animal organisation, e.g., ciliated epithelium (v. Dungern), spermatozoa (Landsteiner, Metchnikoff, Moxter), kidney cells, and leucocytes. These "Antikorper" are also of a complex nature. They obey the already described law of elective absorption, and their origin is in keeping with the "side-chain" theory. It is to be hoped that such immunisations as these, which are of great theoretical interest, may also come to b available for therapeutic application. The idea has already been mooted by v. Dungern, of attacking epithelial new formations, partictularly carcinoma, by means of specific "antiepithelial sera," and Metchnikoff has expressed the somewhat bold hope of being able to delay old age by means of a serum directed against phagocytes (macrophages). But even if in the immediate future no great practical success is attained, we must remember that we are only at the very beginning of a rational investigation of properties of cells which hitherto have been far too lightly regarded. |
Importance of studying both humoral and cellular aspects of immunity
The sifting of the material obtained by observation is rendered more difficult by the occurrence under normal conditions of a great number of quite unlooked for bodies furnished with haptophore groups and arising from diverse organs, and which we may designate collectively as haptines. It is to be expected that the study of these haptines will not only throw light on the more minute details of cellular metabolism, but also prove fruitful in the fields of pathology and therapeutics. By the fact that we can cause the individual haptines of the cells to pass out into the blood serum by a process of specific immunisation, it becomes possible in the test-tube to analyse more accurately the mode of operation of their binding groups than is possible in the case of the complicated conditions which present themselves in the animal body. The importance, for the study of immunity, of considering the circumstances from a purely cellular standpoint is evident from all that I have said. I trust, my lords and gentlemen, that from what I have said you may have obtained the impression, to allude again to my quotation from Bacon, that we no longer find ourselves lost on a boundless sea, but that we have already caught a distinct glimpse of the land which we hope, nay, which we expect, will yield rich treasures for biology and therapeutics. I desire to express my indebtedness to Dr. E. F. Bashford, McCosh Scholar of the University of Edinburgh, now working with me in my Institute, for his kindness in undertaking the translation of my lecture into English, a task to which he has devoted much time and trouble. |
![]()
Some additional viewpoints on Ehrlich's work
Lederberg, J, (1988) " Ontogeny of the clonal selection theory of antibody formation. Reflections on Darwin and Ehrlich. Annals New York Academy of Sciences 546, 175-182.
Forsdyke, D. R. (1995) "The Origins of the Clonal Selection Theory of Immunity" (Click Here)
Klein, J. (1995) "Ehrlich and Darwin. Homochauvinism in immunology. Immunology and Cell Biology 73, 103-108.
Silverstein, A. M. (1996) "Paul Ehrlich: The founding of pediatric immunology". Cellular Immunology174, 1-6.
![]()
Further Acknowledgements for these Immunology Pages
David Talmage kindly provided photographs. Photographs of some other protagonists may be viewed in the image collection of the History of Medicine Division of the National Library of Medicine, USA. Click here for Image Collection. The MHC figure was, with permission, from Colin Hewitt's fine web-page (Click here).
![]()
Other Web Sites on Theoretical Immunology
FORUM. A forum for discussion of matters immunological is the internet newsgroup Bionet.immunology, which was started in the early 1990s by Benny Shomer (Israel) and Donald Forsdyke (Canada).
![]()
The Lymphocyte in Graft and Tumour Rejection (1926)
Positive Selection (1975-2012)
Codon Choice: Antibody, T-cell Receptor & MHC Genes (1986)
Intracellular Self/Not-Self Discrimination
Heat Shock Proteins and Immunity (1985)
Early Evolution of MHC Polymorphism (1991)
Two Signal Model of Self/Not-self Immune Discrimination (1992)
Entropy-Driven Protein Self-Aggregation (1995)
Aggregation-Induced Danger Signals and Cytotoxic T Cells (1999)
Video of this paper with debate between Srivastava and Forsdyke (2015 You Tube version; 53 min) (Vimeo version here)
Chaperone Mode-Switches and Double-Stranded RNA (2000)
Polymorphism and Intracellular Self/Not-Self Discrimination (2001)
Immunity as a Function of the Unicellular State (2002)
Heat-Shock Response and Intracellular Self/Not-Self Discrimination (2005)
X-Chromosome and Intracellular Self/Not-Self Discrimination (2009)
Immunology (1955-1975) reviewed 2012
Lymphocyte Repertoire Selection (2015) Imm. Cell Biol 93, 297-304
![]()
Return to Theoretical Immunology index
![]()
This page was established circa 1999 and last edited 14 April, 2019 by Donald Forsdyke
WWW page access counterSince 30th March 1999

