INTRACELLULAR SELF/NOT-SELF DISCRIMINATION
The following papers provide a mechanism, operating at the protein level, by which an organism might distinguish self from not-self intracellularly, thus leading to the presentation of peptides in association with MHC proteins, so recording a not-self presence in the cell. The intracellular alarm signal which triggers such display would be the presence of a virus protein, or a mutated "self" protein. These would form homoaggregates (and also heteroaggregates with unmutated "self" proteins) which would then trigger protein degradation and peptide formation for MHC. Double-stranded RNA is also implicated. It is not until (and if) this alarm rings that an intracellular protein is labelled as "not-self" and peptides presented. Thus, until this moment, the body distinguishes mainly extracellular self from not-self. This idea has become of increasing interest to cellular immunologists (see Matzinger's "danger" hypothesis 1994. Annu. Rev. Immunol. 12, 991-1045). But a word of warning to researchers. Although you may find the ideas appealing, be cautioned. If you mention them in your next grant application the reviewers may not share your enthusiasm (see web page on Peer Review; Click here). Hint: the first time through you might find it easier to go to the 1995 paper, or read "A Digression on Prions". (Click Here) |
![]()
Heat Shock Proteins and Immunity (1985)
Early Evolution of MHC Polymorphism (1991)
Two Signal Model of Self/Not-self Immune Discrimination: An Update (1992)
Entropy-Driven Protein Self-Aggregation (1995)
Aggregation-Induced Danger Signals (1999)
Chaperone Mode-Switches and Double-Stranded RNA (2000)
Role of Polymorphism in Self/Not-self Discrimination (2001)
Immunity as a Function of the Unicellular State (2002)
Heat Shock Response and Self/Not-Self Discrimination (2005)
X-Chromosome Reactivation and Self/Not-Self Discrimination (2009)
![]()
Heat Shock Proteins and Immunity.
In the early 1980s, prior to the discovery of antigen presentation to T cells as MHC-peptide complexes, it was difficult to link the various phenomena associated with heat shock proteins to immunity. The ideas outlined below were in place by 1985 as the MHC-peptide story began to unfold (Townsend et al. 1985; Babbitt et al. 1985), and a more satisfying (though not necessarily correct) linkage began to emerge (Forsdyke, 1991; Click Here) |
![]()
Heat Shock Proteins Defend Against Intracellular Pathogens: a Non-Immunological Basis for Self/Not-Self Discrimination
D. R. Forsdyke, LETTER TO THE EDITOR 1985
[With copyright
permission from Academic Press]
J. Theoretical Biology 115, 471-473 (Received in revised form 7 January 1985)
When cells of a variety of species are subjected to certain metabolic stresses, such as an increase of temperature, there is a decrease in normal protein synthesis and the induction of a set of several specific proteins which accumulate intracellularly (Schlesinger, Ashburner & Tissieres, 1982). The adaptive value of this "heat shock" response is unknown. In a previous communication it was suggested that a set of extracellular proteins which accumulate in certain clinical conditions, act to cause the aggregation of extracellular pathogens in regions of stagnant blood flow (e.g. the sinusoids of the spleen; Forsdyke & Ford, 1983a). By analogy, it was suggested that heat shock proteins might cause a similar aggregation of intracellular pathogens. There have been reports that heat shock proteins are indeed induced when cells are infected with viruses (Nevins, 1982; Collins & Hightower, 1982; Khandjian & Turler, 1983; LaThangue et al., 1984). I here outline more fully the hypothesis that heat shock proteins create conditions which promote the aggregation ("phase separation") of particles of an intracellular pathogen when particle concentration exceeds a critical value. To eliminate a pathogen, the pathogen must first be recognized as different from the host. Pathogens differ from their hosts in one or more of the following respects.
A mechanism by which a host could exploit the first two of these differences to defend itself against a pathogen has been discussed elsewhere (Forsdyke, 1969, 1975). I here draw attention to a mechanism by which a host could exploit the third difference. When molecules in solution reach a critical concentration, it becomes energetically more favourable for them to aggregate (form networks, crystallize, precipitate) than to remain in simple solution. When certain lipids do this ordered structures (membranes) result: the process is referred to as a lyotropic (concentration-dependent) phase transition. Without any obvious change in solution appearance, some high molecular weight polymers (e.g. Ficoll, polyvinylpyrrolidone, dextran sulphate, heated albumin) form networks in solution at high concentrations (Ogston & Woods, 1954; Comper & Laurent, 1978; Wahl, Stern & Stark, 1979; Minton, 1981; Fulton, 1982). This becomes apparent when other polymers or particles (e.g. erythrocytes) are added. The added polymers or particles are excluded from the phase of the original polymer molecules ("excluded volume effect") and become locally concentrated. This increases the chemical activity of the added polymers or particles, which may aggregate and form ordered structures.
In this light, I propose that the production of heat-shock proteins following infection with an intracellular pathogen (Cosgrove & Brown, 1983) would create intracellular conditions which would promote the phase-separation and aggregation of particles of the pathogen. This aggregation would occur at a far lower concentration of the particles than the concentration required for aggregation in the absence of the heat-shock proteins. The hypothesis predicts that some heat shock proteins should form networks which would exclude other polymers and particles from the solution phase. Work in the author's laboratory was supported by the Physicians' Services Incorporated Foundation, the Leukaemia Research Fund of Toronto, and the Canadian Medical Research Council. REFERENCES Collins, P. L. & Hightower, L. E. (1982) J.
Virol. 44, 703. |
![]()
Early Evolution of MHC Polymorphism
By D. R. FORSDYKE
Journal of Theoretical Biology (1991) 150, 451-456
(Received on 1 May 1990, Accepted on 3 January 1991)
(Reproduced with copyright permission from Academic Press.)
1. Introduction3. Multicellularity Provokes Acceleration of the Arms Race
4. Simplicity Precedes Complexity. Positive Selection
5. Intracellular Distinction between Self and Not-self
6. Positive Selection in Different Environments. Cell Lineages
Abstract. There is unwarranted satisfaction with the view that MHC polymorphism evolved because there was a selective advantage in having a variety of MHC proteins to bind a variety of peptide subsets for presentation to T cells. While this may, in part, explain its maintainance, polymorphism may have evolved initially to reject foreign virus "grafts". The possession of similar membranes promotes aggregation between "like" cells, but it also promotes aggregation between the cells and viruses which retain membrane components of their previous host. The selection pressure afforded by hostile virus "grafts" would favour cells which developed polymorphic membrane components (since "like" will not aggregate with "not-like"). This polymorphism would have evolved before the appearance of multicellular organisms. Thus, the evolution of modern immune systems would have been imposed upon pre-existing polymorphic systems. A path this evolution may have taken involves the development of mechanisms for intracellular distinction between self and not-self. |
|
The hypotheses that the education of potential immunologically competent cells requires both negative selection (Burnet, 1959) and positive selection (Forsdyke, 1975) have found considerable experimental support (reviewed in Schwartz, 1989). However, the relationship of these processes to the phenomenon of MHC polymorphism is poorly understood. It is argued that, although histocompatibility antigens were first recognized through tissue grafting experiments, such grafting does not normally occur in nature. Thus, MHC polymorphism could not have evolved in response to a selection pressure generated by the need to reject foreign grafts. So we must look elsewhere for an explanation for the origin of MHC polymorphism. This view is widely disseminated in textbooks and popular science journals (Marrack & Kappler, 1986; Hopkins, 1987; Grey et al, 1989; Raff, 1989). The discovery of MHC-restricted immune responsiveness (Zinkernagel & Doherty, 1974) and the role of MHC molecules in antigen presentation (Grey et al, 1989) provided a convenient explanation for MHC polymorphism. Extensive MHC polymorphism means that most individuals are heterozygotes. Since all potentially immunogenic peptides cannot bind to one MHC gene product (Grey et al, 1989), having two MHC gene products doubles the chance that an individual will be able to assemble a functional MHC-peptide complex ("heterozygote advantage"; "overdominant selection"; Hughes & Nei, 1989). However, this factor-of-two advantage does not seem particularly compelling. A doubling of the number of MHC loci would achieve the same result. Those of the "group selectionist" school would argue for a much greater advantage at the population level. Thus, a group which had MHC polymorphism would be likely to have at least some members who would have a MHC protein able to form a functional complex with a peptide from a virulent pathogen. However, the general validity of group selectionist arguments has been questioned (Dawkins, 1976). There would be no need to look to explanations such as the above if graft transplantation were an "experiment of nature". Actually, such experiments have probably been going on since life first evolved. When a virus passes from one host cell to another it may transfer some of the cell membrane components of its former host. These may include MHC proteins (Hecht & Summers, 1976; Bubbers & Lilly, 1977; Gelderblom et al., 1987). Such a virus is, in essence, a graft. Individuals highly polymorphic for a membrane component transferred by the virus are likely to reject the virus "graft" as foreign, just as they would a cellular allograft. This rejection response could have evolved before the evolution of the immune response as we know it today. The membranes of the "survival machines" within which the early "replicators" (RNA, DNA) encased themselves (Dawkins, 1976), are initially likely to have been quite similar to each other. We and others have shown that, in an appropriate environment, it can be thermodynamically favourable for cells with similar surface membranes to aggregate preferentially, "like with like" (Forsdyke & Ford, 1983a,b; Armstrong, 1989). Similarly, a nucleic acid fragment (selfish gene/virus) could readily transfer by budding from its cell of origin and, enveloped in the cell membrane of its former host, aggregating with a "like" target cell (Fig. 1a). |
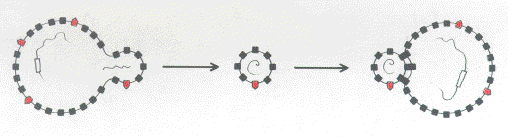
FIG. 1a. Early evolution of virus-cell interactions. (a) A virus enveloped in the membrane of its former host is able to bind to a "like" target cell possessing similar membrane characteristics. Host and virus genomes are represented by wavy lines. The rectangle in the host genome represents a gene encoding a minor host surface component. The latter is represented by open red boxes embedded in the membrane with "caps" pointing to the exterior. |
|
In this circumstance, a cell which had evolved prototypic MHC proteins with some degree of polymorphism, would be better prepared to maintain the integrity of its own nucleic acid by preventing invasion by a foreign nucleic acid (since "like" does not aggregate with "not-like"). MHC proteins could have evolved as a defence against viruses before the evolution of multicellular organisms. The evolutionary pressure to generate extensive polymorphism would have been high, since the greater the polymorphism, the less likely would be a virus to encounter a "like" cell. 3. Multicellularity Provokes Acceleration of the Arms Race However, multicellular organisms, each cell of which would bear "like" polymorphic antigens, would provide a fertile soil for viruses to spread between cells. Thus, the evolution of the multicellular state should be accompanied by a strong pressure for the evolution of immune systems. This evolution would be imposed upon, and might incorporate and adapt, a pre-existing polymorphic MHC system. |
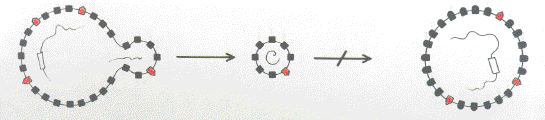
FIG 1b. Polymorphism of membrane components (represented on the left by closed black boxes and on the right by closed black boxes with round caps) prevents viral attachment by the low affinity like-with-like mechanism. |
|
As long as they depended upon the like-with-like mechanism to associate with their target cell membrane, virus envelopes would have been indistinguishable from self. However, at an early stage of their evolution, balked by the development of MHC-polymorphism, viruses would have developed specific coat proteins capable of recognizing non-polymorphic host cell surface proteins as receptors for virus attachment. Steps in developing this capacity might have included:
|
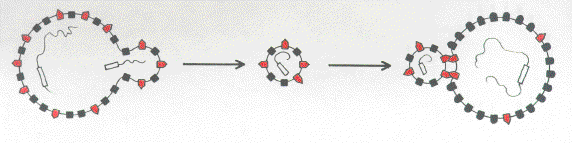
FIG 1c. A virus incorporates the host gene (open rectangle) encoding the minor membrane component (red) and directs increased synthesis of this component. This permits low efficiency encounters with its target cell by the like-with-like mechanism. |

FIG 1d. Mutation of the viral copy of the host gene (shaded rectangle) to generate a surface ligand with high affinity for the target cell receptor. |
|
Thus, the pre-existing like-with-like affinity of host molecules presenting non-polymorphic surface determinants would have been enhanced by mutations of the viral copy of the same gene. The virus coat protein ligand and its host cell receptor would have evolved from a common source, a prototypic non-polymorphic host cell receptor. In entering an "arms race" with its host (Dawkins, 1976), the virus would have "shown its colours" (i.e. acquired a virus-specific surface protein), thus paving the way for specific recognition by a prototypic host immune system. Initially this might have taken the form of the release of soluble decoy receptors from target cells (Ehrlich, 1900). With tissue differentiation, certain cells would specialize in generating decoy receptors. A local tendency to hypermutate would generate a decoy receptor (prototypic antibody) repertoire which might protect organisms against a range of viruses. This would require the coevolution of extracellular mechanisms for distinguishing self from not-self, as discussed previously (Forsdyke, 1968, 1969, 1975). In such a frame-work the immune system as we know it today could have evolved. 4. Simplicity Precedes Complexity. Positive Selection It is possible that one prototypic antibody locus (with multiple V region genes) would have had the opportunity to develop evolutionarily before duplication to generate other loci. A simple model has been considered elsewhere, perhaps corresponding to an organism which once existed (or may even still exist). The model organism had a single hypervariable locus and could have mounted both cell-mediated and humoral immune responses (Forsdyke, 1968, 1969, 1975). The model proposed that enhanced immunological reactivity against "near-self " MHC antigens ("alloaggression") is not encoded in the germ line as suggested by Jerne (1971), but requires a phase of positive selection of the immunological repertoire (Forsdyke, 1975). MHC proteins and other membrane components (Bluestone et al, 1988; Porcelli et al, 1989; Vidovik et al, 1989) would play a major role in the shaping of this repertoire. It was also pointed out that expanded clones of anti-near-self immunologically competent cells would present a barrier opposing the progressive mutation of molecules of a pathogen towards forms indistinguishable from host molecules ("molecular mimicry"). 5. Intracellular Distinction between Self and Not-self It is argued (Bevan, 1989) that:
Recently it was stated (Kourilsky & Claverie, 1989) that the:
Thus, it is envisaged (Bevan, 1989) that:
However, absence of evidence is not evidence of absence. Absence of a known mechanism for intracellular self/non-self distinction does not mean that such a mechanism does not exist. Indeed, the principle of intracellular self/non-self distinction was firmly established with the discovery of bacterial DNA restriction- modification systems (Arber & Linn, 1969). A mechanism for distinction at the protein level, based on concentration-dependent (lyotropic) phase transitions, has been proposed (Forsdyke, 1985). Proteins which exceed their solubility limits in the crowded intracellular environment will aggregate and mark themselves as different from other proteins (Marston, 1986). The synthesis and breakdown of viral proteins would not be so tightly regulated as in the case of host cell proteins. After all, the raison d'tre of a virus is to increase in number. Changes in environmental conditions (e.g. heat shock), which might favour the aggregation of self proteins, would be dealt with by special mechanisms, such as the activation of chaperonin-like proteins which would reverse aggregation (Fischer & Schmid, 1990). Having been marked as different, foreign aggregates would be degraded, first to peptides and then to amino acids. This level of coping with foreign material could be achieved by unicellular organisms. Association of foreign peptide fragments with prototypic polymorphic MHC proteins en route to the cell surface would not seem advantageous at the unicellular stage of evolution. However, in multicellular organisms, the modified, "near-self ", MHC antigenic determinants so created would be recognized by host immunologically competent cells which would already have been educated, by mechanisms such as outlined previously (Forsdyke, 1975), to respond against near-self. By associating peptide fragments with MHC determinants, the organism would merely be taking advantage of the pre-existence of a large population of immunologically competent cells reactive with near-self. A cell displaying modified MHC proteins would have become labelled for destruction by immunologically competent cells or their products, a feature of considerable selective advantage for the organism. The pre-existing polymorphism of MHC proteins would increase the chance that a successful MHC-peptide complex would be formed and, in the continuing evolutionary arms race with pathogens, modifications of MHC proteins might have occurred to enhance their recognition of sets of structural features shared by many peptides. Since the critical self-vs.-non-self decision had already been made intracellularly, it would be sufficient for the peptide-MHC protein complex to be recognizable as near-self. This might involve recognition by T cell receptors both of conformational changes in MHC molecules and of features of the associated peptide. There would be no necessity for a stage in the education of immunologically competent cells in which cells reactive with complexes of MHC proteins and self-peptides derived from the organism's intracellular proteins would be eliminated (Berg et al. 1989). This would then leave open to the organism the option of destroying cells in which the regulation of the intracellular concentration of one of its own proteins had become disordered. 6. Positive Selection in Different Environments. Cell Lineages Duplication of hypervariable loci and segregation of their expression to different tissue compartments would provide opportunity for exposure of different populations of potential immunologically competent cells to different spectra of antigens during the phase of positive selection. Thus, cell lineages (e.g. prototypic T and B lineages) might have arisen. This would provide the opportunity for the evolution of a division of labour. One lineage would be concerned primarily with cell-mediated responses and another lineage with humoral responses.[For further discussion of lineages, please see Forsdyke (1992) J. theor. Biol. 154, 109-118.] |
REFERENCES
ARBER, W. & LINN, S. (1969). Ann. Rev.
Biochem. 38,467-500.
ARMSTRONG, P. B. (1989). Crit. Rev. Biochem. 24, 119-149.
BERG, L. J., PULLEN, A. M., FAZEKAS DE ST. GROTH, B., MATHIS, D., BENOIST, C. & DAVIS,
M. M. (1989). Cell 58, 1035-1046.
BEVAN, M. J. (1989). Nature, Lond. 342, 478-479.
BLUESTONE, J. A., CRON, R. Q., COTTERMAN, M., HOULDEN, B. A. & MATIS, L. A. (1988).
J.
expl. Med. 168, 1899-1916.
BUBBERS, J. E. & LILLY, F. (1977). Nature, Lond. 266, 458-459.
BURNET, F. M. (1959). The Clonal Selection Theory of
Acquired Immunity. Cambridge: Cambridge University Press.
DAWKINS, R. (1976). The Selfish Gene.
Oxford: Oxford University Press.
EHRLICH, P. (1900). Proc. Royal. Soc. (London) 66,424-435.
FISCHER, G. & SCHMID, F. X. (1990). Biochemistry 29, 2205-2212.
FORSDYKE, D. R. (1968). Lancet 1, 281-283.
FORSDYKE, D. R. (1969). J. theor. Biol 25, 173-185.
FORSDYKE, D. R. (1975). J. theor. Biol 52, 187-198.
FORSDYKE, D. R. (1985). J. theor. Biol 115, 471-473.
FORSDYKE, D. R. & FORD, P. (1983a). J. theor. Biol. 103, 467-472.
FORSDYKE, D. R. & FORD, P. (1983b). Biochem. J. 214,257-260.
GELDERBLOM. H., REUPKE, H., WINKEL, T., KUNZE, R. & PAULI, G. (1987). Z Naturforsch.
42c, 1328-1334.
GREY, H. M., SETTE, A. & BUUS, S. (1989). Sci. Am. 261, (5), 56-64.
HECHT, T. T. & SUMMERS, D. F. (1976). J. Virol. 19, 833-845.
HOPKI NS, N. H. (1987). In: Molecular Biology of the Gene,
4th edn Menlo park: Benjamin/Cummings. p. 888.
HUGHES, A. L. & NEI, M. (1989). Proc. natn. Acad. Sci.
U.S.A. 86,958-962.
JERNE, N. (1971). Eur. J. Immunol 1, 1-9.
KOURILSKY, P. & CLAVERIE, J. M. (1989). Cell 56, 327-329.
MARGULIES, D. H. (1989). Nature, Lond. 342, 124-125.
MARRACK, P. & KAPPLER, J. (1986). Sci. Am. 254 (2), 36-45.
MARSTON, F. A. 0. (1986). Biochem. J. 240, 1-12.
PORCELLI, S., BRENNER, M. B., GREENSTEIN, J. L., BALK, S. P., TERHORST, C. & BLEICHER,
P. A. (1989). Nature, Lond 341, 447-450.
RAFF, M. (1989). In: Molecular Biology of the Cell,
2nd edn, New York: Garland. p. 1040.
SCHWARTZ, R. H. (1989). Cell 57,1073-1081.
VIDOVIC, D., ROGLIC, M., MCKUNE, K., GUERDER, S., MACKAY, C. & DEMBIC, Z. (1989). Nature,
Lond. 340, 646-649.
ZINKERNAGEL, R. M. & DOHERTY, P. C. (1974). Nature, Lond. 248,701-702.
![]()
End Note on Heterozygote Advantage May 2014
| Borghans et al. 2004 write: "Heterozygote
advantage per se is insufficient to explain the high degree of
polymorphism at the MHC, even in very large host populations.
Host-pathogen coevolution, on the other hand, can easily account for
realistic polymorphisms of even more than 50 alleles per MHC locus." Borghans, Beltman & De Boer (2004) Immunogenetics 55, 732-739. MHC polymorphism under host-pathogen coevolution. |
![]()
Two Signal Model of Self/Not-self Immune Discrimination: An Update
D. R. FORSDYKE
Journal of Theoretical Biology 154, 109-118 (With copyright permission from Academic Press; Received on 25 February 1991, Accepted on 21 May 1991)
1. Introduction
2. Some Principles
3. Prototypic B Cells First
4. Education of Prototypic B Cells
5. Education of Prototypic T Cells
7. T Cell Subsets
8. Intracellular Self/Not-self
Discrimination
9. Two Gates for T Cells
From a consideration of the evolution of the first immune systems in simple organisms, it is proposed that mechanisms for self/not-self discrimination evolved at an early stage of evolution in prototypic B cells, prior to separation of the B cell and T cell lineages. After passing an MHC protein-limited self/not-self discrimination gate, T cells depend for fine self/not-self discrimination either on B cells (extracellular discrimination) or on cells harbouring pathogens (intracellular discrimination). Full activation of B cells needs interaction with T cells, but this evolved late and is not part of the self/not-self decision. |
|
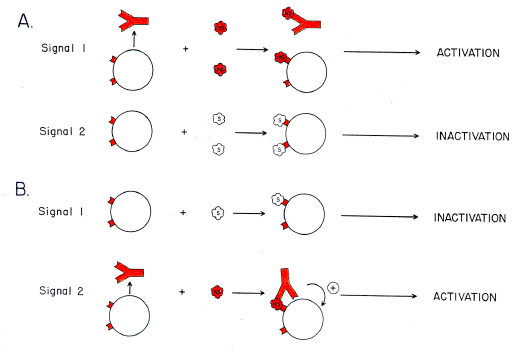
Immunologically competent cells (ICCs) are lymphocytes of either the B or T cell lineage. Burnet (1959) and Lederberg (1959) originally proposed that generation of ICCs involves an early time-window during which cells bearing antibody-like receptors are destroyed on encountering specific self antigenic determinants which react with these receptors. Cells surviving this encounter pass through the window and respond immunologically if they then encounter specific not-self antigenic determinants. Thus cells receive one signal from antigen, but respond differentially depending on their physiological state. These ideas were subsequently elaborated to accommodate the ability of high antigen concentrations to inhibit the immune response in adults (Mitchison, 1964), and the possibility that cells which have passed the window might mutate to react to self antigenic determinants. Extending earlier work of Medawar (1963) and Nossal (1965), I proposed that antigenic determinants are capable of giving either of two signals to specific lymphocytes at any stage of their development [Forsdyke, 1966, 1968, 1969; Fig. l(a)]. |
|
|
FIG. 1. Two signal models for lymphocyte activation and inactivation. Elements shown are cells with antibody-like receptors, free antibody and antigen. The latter may either be self (S) or not-self (NS).
|
|
The effective concentration of an antigenic determinant depends, not so much on its absolute concentration within the body, but on the ratio of the concentration of antigen to the concentration of available antigen-binding activity. In an unimmunized organism the latter is largely provided by natural antibody. At an early stage of immunological maturation there is little specific extracellular antigen-binding activity. Unbuffered by this activity, a specific lymphocyte encounters a high effective antigen concentration and is destroyed (signal 2). Later the concentration of specific antibody increases so that effective antigen concentrations tend to be lower. The signal given to a specific lymphocyte then induces an immune response (signal 1). Thus, the outcome of reaction of a lymphocyte with antigenic determinants is critically dependent on a second antigen-recognizing activity, natural antibody, the concentration of which is time-dependent [Fig. l(a)]. The model is "two signal" in another sense. Signal I requires limited receptor occupancy (minimum one signal), whereas signal 2 requires reaction with a second receptor in close proximity to the first (two signals). An alternative formulation, closer to earlier ideas (Medawar, 1963; Nossal, 1965) was offered by Bretcher & Cohn (1968, 1970; reviewed by Cohn in Langman, 1989). They proposed that the second antigen-recognizing activity actually delivers a positive signal to a specific lymphocyte [Fig. 1(b)]. This "helper" activity, initially envisioned as free antibody and later as antibody synthesized and borne by another lymphocyte, is essential for an immune response. In the absence of the signal from the second antigen-recognizing activity there is cell destruction. Focusing of the latter activity to a lymphocyte requires that an antigen present at least two antigenic determinants. Under this model inactivation occurs slowly and is by default (failure to receive a help signal). Under my two signal model inactivation is an active process, which in some circumstances may involve complement (for references see Forsdyke, 1975, 1977, 1980). Cohn and coworkers have recently argued that their model is consistent with modern concepts in immunology (Langman, 1989; Cohn & Langman, 1990a, b, c). Here a similar case is made for my model. Some aspects of this work are discussed more fully elsewhere (Forsdyke, 1985, 1991).
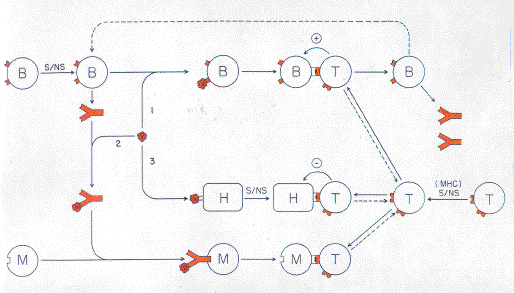
The result of a long evolutionary process, modern vertebrate immune systems appear very complex (Burnet, 1978; Marchalonis & Schluter, 1989). However, all immune systems have basic features which would have evolved at an early stage. It seems likely that organisms with one set of antibody variable (V) region genes would have evolved and have achieved some degree of immunological sophistication before being superseded by organisms with two sets of V region genes. A prototypic immune system with one set of V region genes might have displayed all basic system features, such as discrimination between self and not-self and the ability to mount both cellmediated and humoral immune responses (Forsdyke, 1969). Duplication of the set of V-region genes would have produced an initial redundancy followed by a division of labour so that one set of V-regions would have become specialized for cell-mediated responses and another for humoral responses. There would have been an interplay between the cell-mediated and humoral response systems, so that each would have evolved in the context of the other. Similarly there would have been an evolutionary interplay with other coevolving systems, such as those concerned with metabolic regulation and with maintaining cell population-size homeostasis (Rocha et al., 1989). Along similar lines, it seems likely that one MHC protein system would have evolved before a second (Lawlor et al., 1990). The second system would have evolved in an organism already possessing the first system. There would have been an evolutionary interplay between the two systems. Prototypic ICCs are held to have properties of both B cells and T cells. They are generated in a "mutant breeding organ" and undergo a subsequent "education" such that high specificity anti-self cells are destroyed (negative selection) and low specificity anti-self cells accumulate (positive selection; Forsdyke, 1975). Duplication of the prototype V-region set would facilitate the development of independent systems of B cells and T cells. The steps required to develop a mature B cell would appear less complex than those required to develop a mature T cell. Whereas progenitors of both B cells and T cells arise in the bone marrow, T cell progenitors must migrate to the thymus to become ICCs. Thus, it is postulated that a prototypic mutant breeding organ was bone marrow-like and, in this respect, prototypic ICCs were more like B than T cells. In the bone marrow positive selection could be influenced by a wide range of self antigens and hence might not be well-developed for any particular antigen. However, sequestration of potential ICCs in a compartment where they could be selectively exposed only to a narrow range of self antigens would result in the positive selection of a restricted set of low specificity anti-self cells. The selective advantage of accumulating ICCs of low specificity for self MHC antigens has been discussed previously (Forsdyke, 1975, 1991). Thus, the evolution of a compartment where potential ICCs could be exposed selectively to self MHC antigens would be favoured. The result of this process would be the thymus of modern vertebrates. The process would involve evolution of a germ line V region gene set with an enhanced probability of efficiently generating variable regions capable of low specificity reaction with self MHC. Duplication of the prototypic V region set would have released a set of V region genes from prototypic B cell constraints (marrow-based) and facilitated the evolution of this set of V region genes dedicated to prototypic T cells (thymus-based). 4. Education of Prototypic B Cells After association of a particular prototypic variable region gene with a prototypic constant region gene, the corresponding IgM-like immunoglobulin would be assembled in prototypic B cells and first displayed as a receptor at the cell surface. At this unique time-point and in the absence of the corresponding free antibody, the effective concentration of self antigenic determinants (even if present at low absolute concentrations) would be high. By mechanisms such as previously outlined (Forsdyke, 1966, 1968, 1969), reaction of a potential ICC with high concentrations of such determinants would result in cell destruction. Failing this, and following positive selection by a wide range of self antigenic determinants (Forsdyke, 1975), immunoglobulin would be secreted generating a pool of natural antibody capable of buffering the cell against future reaction with excess antigen. In these circumstances, the effective concentration of most foreign antigens would be low and reaction with the cell would trigger enhanced secretion of the appropriate antibody. 5. Education of Prototypic T Cells Within the thymic compartment, prototypic T cells would be separated from the antibody secreted by prototypic B cells. The T cells would be exposed to a limited spectrum of self-antigens, primarily MHC antigens. Cells bearing receptors of high specificity for these antigens would be destroyed (negative selection). Cells with receptors of low specificity would accumulate (positive selection). Cells with very low to zero specificity would degenerate (Forsdyke, 1975; Bevan, 1977; Blackman et al., 1990; Boehmer & Kisielow, 1990; Rothenberg.. 1990). Moving through different body compartments (e.g. blood, lymph, lymph-nodes) the selected T cells would be subjected to various degrees of buffering against foreign antigens by antibodies secreted by prototypic B cells. 6. T Cells Permissive for B Cells Let us consider the immune response to a virus of an evolutionarily simple organism with one prototypic MHC locus and two sets of variable region genes (one for prototypic B cells and one for prototypic T cells; Fig. 2). |
|
|
| FIG. 2. Response to an antigen in an evolutionarily simple host organism
in which prototypic MHC genes have not
duplicated.
Various elements shown are a virus antigen (V; coloured red), a prototypic B cell (B), a prototypic T cell (T), a phagocytic cell (M), a host cell (H; bearing a receptor for virus), and free antibody. Dashed lines indicate clonal expansion of a particular cell type in response to an appropriate signal. A prototypic B cell (top left), passes the self/not-self discrimination gate (S/NS) and begins synthesis of natural antibody. A foreign virus has three fates:
|
|
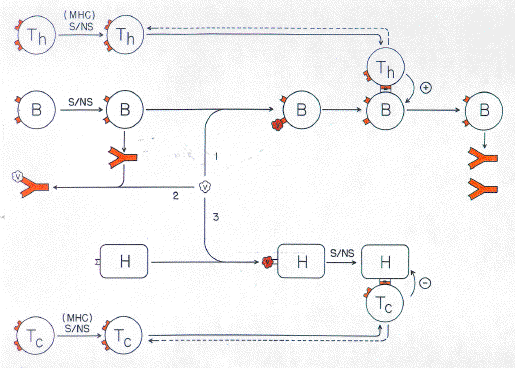
Some viruses react with natural antibody and thus are labelled as not-self. Appropriately opsonized, the viruses attach to and are ingested by phagocytic cells. In either a free or attached state, native virus protein epitopes also react with specific B cells. This satisfies one condition necessary for the B cells to respond by proliferation and enhanced secretion of antibody. Meanwhile, some viruses, having evaded capture by antibody, penetrate their host cell targets and viral proteins are synthesized. By mechanisms such as discussed elsewhere (Forsdyke, 1985, 1991), the viral proteins are recognized as foreign intracellularly. They are then degraded and peptide fragments displayed at the host cell surface in association with MHC protein (Townsend & Bodmer, 1989). This complex acts as a recognition site stimulating prototypic T cells with specific receptors to proliferate and/or destroy the host cell. Within phagocytic cells viral proteins follow a degradation pathway similar to that within host target cells, except that entry into the pathway does not depend upon intracellular self/not-self discrimination. The peptides eventually appear at the cell surface in association with MHC protein (Fig. 2). The MHC protein-peptide complex could, in principle, be recognized by either prototypic B cells or prototypic T cells bearing receptors of the appropriate specificity. However, because of thymic positive selection there would be more potentially reactive T cells. Furthermore, a T cell specific prototypic CD4/CD8-like molecule would evolve to enhance interaction of the T-cell receptors with the MHC protein-peptide complex. Prototypic B cells, like host target cells and phagocytic cells, incorporate and process viruses and display peptides at their surfaces complexed with MHC protein (Fig. 2). Complexes are recognized by prototypic T cells which are activated to further proliferate and/or deliver, not a cytotoxic signal, but a specific "help" signal, possibly involving a short-lived cytokine. This satisifes a second and final condition for B cell activation, but is not related to the process of self/not-self discrimination. How did this activation of prototypic B cells (capable eventually of secreting small diffusible IgG-like molecules) become dependent on a reaction with prototypic T cells? There are conditions underwhich a humoral response can exclude a cell-mediated response (for references see Forsdyke, 1969). If it were more important to mount a T cell response than a B cell response, then T cell control of the B cell response could evolve. For example, a virus-infected host cell might display native viral epitopes at the cell surface (for references see Forsdyke, 1991). Reaction of antibody with these epitopes might hinder concomitant reaction of T cells with MHC protein-peptide complexes borne by the same host cell. Indeed, to correct for the possibility of B cell activation in the absence of T cell help, there might evolve a mechanism for T cells in some circumstances actually to deliver a negative signal to activated B cells (Shinohara et al., 1988). A prototypic T cell activated by a peptide-MHC complex proliferates thus expanding clone size and creating daughter cells (immunological memory; Fig. 2). Any cell displaying the same MHC protein-peptide complex is a potential target of the daughter cells. Possessing both cytotoxic and helper capacity, these cells have to determine whether the target is a member of the phagocytic and B cell (lymphoreticular) lineages (requiring help) or is a cell hosting a pathogen (requiring destruction). The latter cells do not normally respond to the local help signal. Cells of lymphoreticular lineages respond to the local help signal and would have to evolve some mechanism to resist the cytotoxic signal. However, this would leave lymphoreticular cells vulnerable to intracellular pathogens, so there should be a selection pressure favouring the evolution of precise discrimination between help and cytotoxic signals. Prototypic T cells would have both helper and cytotoxic functions. In the course of evolution duplication of appropriate genes would create an opportunity for T cell subsets to arise with an appropriate division of labour. Thus, T cells recognizing MHC protein peptide complexes on virus-infected host cells would be predominantly cytotoxic. T cells recognizing MHC protein-peptide complexes only on lymphoreticular cell lineages would specialize in helper functions. This division of labour would involve duplication of prototypic MHC loci and duplication of the T cell specific prototypic CD4/CD8-like locus. Thus, one set of T cells (prototypic CD4 bearing) would arise with specificity for peptides presented by products of one set of MHC genes (MHC class II prototypes). These cells would be primarily concerned with giving permissive signals to appropriate MHC class II protein-bearing cells. Another set of T cells (prototypic CD8 bearing) would arise with specificity for peptides presented by products of another set of MHC genes (MHC class I prototypes). These cells would be primarily concerned with the destruction of appropriate MHC class I protein-bearing cells (Fig. 3). Cells of the lymphoreticular lineage, bearing both classes of MHC molecule, would be potential targets of both types of T cells. The evolution of a dependence of T cells-on further signals (e.g. interleukin-1) will not be considered here. |
|
|
FIG. 3. Response to an antigen in an evolutionarily more complex host organism in which two distinct prototypic T cell subsets have arisen (Th,Tc), and the prototypic MHC locus and prototypic CD4/CD8-encoding locus have duplicated. Other elements shown are virus antigen (V), a prototypic B cell (B), a host cell target (H) and free antibody. The role of phagocytic cells is not shown. Dashed lines indicate cell activation to proliferate (clonal expansion). Full self/not-self discrimination (S/NS) operates extracellularly at the level of B cells and intracellularly at the level of host cells. MHC protein-limited self/notself discrimination occurs extracellularly at the level of T cells. The three fates of virus antigen are similar to those shown in Fig. 2 except that separate T cell lineages deliver the permissive signal for the B cell immune response (Th) and the negative signal for host cell destruction (Tc). |
|
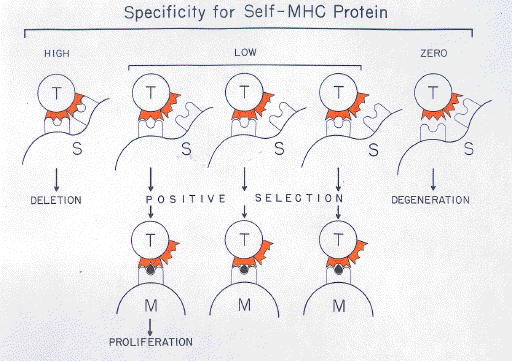
8. Intracellular Self/Not-self Discrimination The postulated events leading to the activation of cytotoxic T cells include an intracellular mechanism for self/not-self discrimination, elements of which may have existed prior to the evolution of multicellular organisms (Forsdyke, 1985, 1991). This mechanism would be of a "housekeeping" nature and would be present in all cells including the thymic stromal cells which display the MHC proteins involved in positive and negative selection. If intracellular self/not-self discrimination by these cells is error-free then, in the absence of intra-thymic not-self antigens, thymic MHC proteins should be empty and not associated even with "nominal" peptide (Berg et al., 1989; Howard, 1990). However, in principle, there seems to be no reason why some self peptides should not declare themselves as such by associating with MHC protein during the stage of thymic education. Included in this category might be small physiologically active circulating peptides which might diffuse into the thymus. Furthermore, should the intracellular self/not-self discrimination mechanism be error-prone with respect to certain self proteins, this would be the time to delete the precursors of T cells capable of reacting with MHC proteins containing peptide fragments derived from these self proteins. For logistic and other reasons (Forsdyke, 1991), it is not envisaged that all possible self peptides are displayed during thymic education. Prior screening to eliminate T cells capable of recognizing self-peptide-associated MHC proteins would not eliminate the logistic problem of competition between self and foreign intracellular peptides for presentation with MHC proteins at the cell surface. Some ad hoc intracellular mechanism for discrimination seems imperative (Lefkowits, 1986; Forsdyke, 1985, 1991). There would be intrathymic negative selection of T cells whose receptors show high specifity for MHC protein and any other self components presented within the thymus. There would be positive selection of the corresponding cells bearing low specificity receptors. Thus, some T cells might emerge from the thymus possessing receptors of low specificity to thymic self cell-surface antigens not related to MHC proteins. However, in the absence of CD4/CD8-like bridges linking T cells to monomorphic regions of these surfaces molecules, or some other modification, T cell activation by them seems unlikely. Prototypic T cells would have most resembled cytotoxic T cells (Forsdyke, 1991). With the evolution of the helper T cell lineage there arose a dependence on B cells for recognition of foreignness. The critical event leading to the activation of helper T cells would be recognition of antigen by B cells or by free antibody secreted by these B cells (Figs 2 and 3). The B cells would themselves already have successfully passed the self/non-self discrimination gate. Thus, the extracellular mechanism for self/non-self discrimination postulated for B cells is ultimately also operative for helper T cells. |
|
|
FIG. 4. Education of T cells requires MHC protein-dependent and peptide-dependent gates. Elements shown are T cells (T) bearing receptors of various specificities, thymic stromal cells (S) displaying MHC proteins, and peripheral tissue cells (M; each displaying a peptide from a processed antigen, shown as a filled bullet-like object).
|
In essence, T cells pass through two discrimination gates (Fig. 4). The first gate is extracellular and involves cell surface components within the thymus. Cells passing this gate can survive, but proliferation is limited. Clonal expansion is permitted when the T cell receptor encounters an extra-thymic MHC protein-peptide complex. This latter gate can only exist if the antigen from which the peptide was generated has already been recognized as not-self either intracellularly in a cell which has subjected the antigen to processing in a cytosolic compartment (Townsend & Bodmer, 1989), or extracellularly by B cells or their products. The cytosolic and B cell discrimination mechanisms can be regarded as surrogates for a more direct mechanism of ensuring that T cell receptors are of high specificity for their not-self targets. Thus, B cells and host cells "read" various aspects of an antigen (its concentration and specificity for their receptors) and relay this information to T cells. The latter discriminate primarily on the basis of specificity. I thank R. E. Langman and J. J. Marchalonis for helpful reviews of the manuscript. |
REFERENCES
BERG, L. J., PULLEN, A. M., FAZEKAS DE ST. GROTH, B., MATHIS, D., BENOIST, C.
& DAVIS, M. M. (1989). Cell 58,
1035-1046.
BEVAN, M. J. (1977). Nature, Lond.
269, 417-419,
BLACKMAN, M., KAPPLER, J. & MARRACK, P. (1990). Science
248, 1335 1341.
BOEHMER, VON H. & KISIELOW, P. (1990). Science
248, 1369-1373.
BRETCHER, P. & COHN, M. (1968). Nature, Lond.
220,444-448.
BRETCHER, P. & COHN, M. (1970). Science
169, 1042-1049.
BURNET, F. M. (1959). The Clonal Selection Theory
of Acquired Immunity. Cambridge: Cambridge University Press.
BURNET, F, M. (1978). In Theoretical Immunology
(Bell, G. I., Perelson, A. S. & Pimbley, G. H., eds) pp. 63-85. New York: M.
Dekkar.
COHN, M. & LANGMAN, (1990a). Immunol. Rev.
115, 7-141.
COHN, M. & LANGMAN, R. E, (1990b). Cell.Immunol.
130,213-223.
COHN, M. & LANGMAN, R. E. (1990c). Cell. Immunol. 130, 229 -235.
FORSDYKE, D. R. (1966). In: The Biological Effects
of Phytohaemagglutin (Elves, M. W., ed.) pp. 115-123. Oswestry, U.K.:
R. Jones and A. Hunt Orthopaedic Hospital Management Committee.
FORSDYKE, D. R. (1968). Lancet I,
281-283.
FORSDYKE, D. R. (1969). J. theor. Biol.
25, 173-185.
FORSDYKE, D. R. (1975). J. theor. Biol.
52, 187-198.
FORSDYKE, D. R. (1977). Nature, Lond.
267, 358-360.
FORSDYKE, D. R. (1980). Can. J. Biochem.
58, 1387- 1396.
FORSDYKE, D. R. (1985). J. theor. Biol.
115, 471-473.
FORSDYKE, D. R. (1991). J. theor. Biol.
150, 451-456.
HOWARD, J. C. (1990). Cold Spring Harbour Symp.
quant. Biol. 54, 947-957.
LANGMAN, R. E. (1989). The Immune System.
New York: Academic Press.
LAWLOR, D. A., ZEMMOUR, J., ENNIS, P. D. & PARHAM, P. (1990). Ann. Rev. Immun. 8, 23-63.
LEDERBERG, J. (1959). Science 129,
1649-1653.
LEFKOWITS, I. (1986). Proc. natn. Acad. Sci. U.S.A.
83, 3437-3438.
MARCHALONIS, J. J. & SCHLUTER, S. F. (1989). Dev.
Comp. Immunol. 13,285-301.
MEDAWAR, P. B. (1963). In: Acquired or Natural Immune Tolerance Towards Defined
Protein Antigens (Bussard, A., ed.) pp. 475-480. New York: Academic Press.
MITCHISON, N. A. (1964). Proc. R. Soc. Lond. B.
161, 275-292.
NOSSAL, G. J. V. (1965). Aust. Ann. Med.
14, 321 328.
ROCHA, B., DAUTIGNY, N. & PEREIRA, P. (1989). Eur.
J. Immunol. 19, 905-911.
ROTHENBERG, E. V. (1990). Immun. Today
11, 116-119.
SHINOHARA, N., WATANABE, M., SACHS, D. H. & HOZUMI, N. (1988). Nature, Lond. 336,481-484.
TOWNSEND, S. & BODMER, H. (1989). Ann.Rev.Immunol,
7, 601-624.
![]()
Entropy-Driven Protein Self-Aggregation as the Basis for Self/Not-Self Discrimination in the Crowded Cytosol
By D. R. FORSDYKE
Journal of Biological Systems (1995) 3, 273-287. (Received 14 December 1993)
(With copyright permission from World Scientific Publishing Company)
ABSTRACT
1. The Need for Intracellular Self/Not-Self Discrimination
2. Distinction Between Specific and Collective Protein Functions
3. Self-Aggregation of Red Blood Cells
4. Self-Aggregation of Macromolecules
5. Specificity of Self-Aggregation
6. Proteins Exert a Collective Pressure in the Crowded Cytosol
7. Fine-Tuning of Intracellular Protein Concentrations
8. Intracellular Self/Not-Self Discrimination
9. Heat Shock Proteins Sustain Aggregation Pressure
10. T Cell Memory and Fever
11. Other Compartments and Other Interspecies Relationships
12. Further Implications and Experimental Predictions
End Note 2001a
End Note 2001b
End Note 2006
End Note May 2008
End Note Nov 2008:
End_Note_August_2012
End_Note_May_2014:
End_Note_Feb_2018_Peptides_from_Large_and_Abundant_Proteins
|
ABSTRACT
Cytotoxic T cells recognize cell surface complexes of MHC class I proteins with peptides derived from proteins synthesized within the recognized cell. A mechanism permitting some intracellular discrimination between self proteins and not-self proteins (encoded by a foreign species) would allow the preferential loading of MHC proteins with peptides derived from not-self proteins. This would decrease competition with peptides derived from self proteins and decrease gaps in the T cell repertoire. A possible mechanism has been derived from studies of the specificity of self-aggregation of erythrocytes and of virus coat protein. It is postulated that genes whose products occupy a common cytosol have co-evolved such that product concentrations are fine-tuned to a maximum consistent with avoiding self-aggregation. Cytosolic proteins collectively generate a pressure tending to drive protein molecules into self-aggregates. Each individual protein species both contributes to, and is influenced by, this pressure. The aggregation involves a liberation of bound water and an increase in entropy. The concentrations of proteins encoded by viral genes (not-self) readily exceed the solubility limits imposed by the crowded host cytosol. This leads to their preferential degradation to peptides which associate with MHC proteins. The intracellular and extracellular self/not-self discrimination systems complement each other to ensure that there is no immunological reaction against normal self tissue components. Keywords: Collective protein function, concentration fine-tuning, heat-shock response, evolution. |
|
1. The Need for Intracellular Self/Not-Self Discrimination The maturation of immunologically competent cells involves the deletion or inactivation of cells which react with high specificity with self antigenic determinants. Since the cells first "show their colours" by displaying antigen-specific, outward-facing, receptors at their surfaces, their "education" is regarded as part of an extracellular process of self/not-self discrimination. A cell whose receptors happen to react with some free or cell-borne self-antigenic determinant will either be deleted or inactivated [15, 45]. The principle of intracellular self/not-self discrimination was firmly established with the discovery of bacterial restriction/modification systems [1]. An advantage to higher multicellular organism of having an intracellular self/not-self discrimination system became particularly apparent with the finding that peptides from both self and foreign intracellular proteins can be displayed at the cell surface in association with major histocompatibility complex (MHC) proteins; here they are recognized by immunologically competent cells of the cytotoxic T cell class [43]. |

|
In the absence of a mechanism for distinguishing intracellularly between self peptides or proteins and foreign (e.g., virally-derived) peptides or proteins, cells would have to display all MHC-bound peptides. Hedrick has noted [26] that:
and has suggested that:
In the absence of such a sorting mechanism, an organism would depend solely on prior education of T cells to prevent their being activated by self-peptide-MHC complexes. Several studies indicate that this deletion of potential self-reacting T cells would create numerous gaps in the T cell repertoire, which might adversely affect resistance to infection [42,46,49]. For example, Du Pasquier and coworkers [8] presented evidence that polyploid toad species, while apparently functionally polyploid for most genes, are functionally diploid for MHC genes. That this had occurred over evolutionary time was suggested by the observation that healthy laboratory-made polyploid toads expressed all MHC loci. Since the final T cell repertoire reflects the processes of negative and positive selection [12], every additional functional MHC gene would increase the number of potential anti-self T cells, which would have to be deleted (negative selection). This would decrease the range of foreign proteins to which the surviving positively selected cells could respond. Fitness would be impaired. Individuals who lost excess MHC loci through mutation, would then have a selective advantage so that the modern polyploid survivors would be MHC diploid. As Vidovic and Matzinger succinctly state [49]:
The prior sorting of peptides intracellularly would mean that populations of T cells would exist with the potential to respond against self-peptides should they be inadvertently, or artificially, presented. Thus, Schild and coworkers [46] found that cytotoxic T lymphocytes:
2. Distinction Between Specific and Collective Protein Functions So how could intracellular peptides be sorted? I here review a proposed mechanism of intracellular self/not-self discrimination derived from studies of the specificity of the aggregation of red blood cells [20-22], and tobacco mosaic virus coat protein [31-33]. Although still unproven, the mechanism appears to throw new light on a number of quite disparate phenomena, including fever, the heat shock response, temperature-sensitive mutations and even X chromosome dosage compensation [16-19]. Three basic postulates are that:
3. Self-Aggregation of Red Blood Cells The discovery of this phenomenon has a fascinating early history. The ancient Greeks associated disease with an excess of white "phlegm", one of the four body "humours". To rid the body of phlegm, sick persons were bled. This practice endured for over two millennia. Fahraeus in 1921 [10-11], extending earlier observations of Hewson [27], made a convincing case for the origin of therapeutic bleeding. He noted that in ancient times people would have been bled into earthenware vessels. The clot would have contracted expressing yellow serum. On emptying a vessel the Greeks would first have poured off the serum and then have dislodged the clot. This would have been red in its oxygenated upper part and black in its anaerobic base. Thus, three of the humours would have gained their names, choleric, sanguine, melancholic (yellow, red, black). However, in the case of blood from a sick person, an extra white layer (phlegm) would have been seen above the red part of the clot. What the Greeks did not know was that in freshly drawn stationary blood from a sick person the red cells often aggregate to form rouleaux. These sediment rapidly before a clot can form, leaving a cell-free upper plasma layer. The eventual clot involves not only the red cell mass, but also the upper plasma layer. The clot contracts expressing yellow serum, as with blood from a normal subject, but the dislodged clot now shows three layers (Fig. 1). |
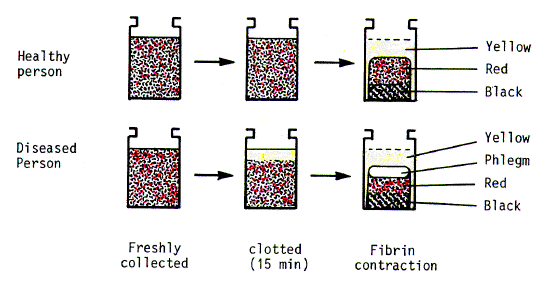
| Fig. 1. Generation of an additional layer
(phlegm) in the contracted clot of blood from a diseased person.
Blood from a healthy person (upper) or diseased person (lower), is either freshly collected (left), freshly clotted (middle), or left overnight for the clot to contract (right). Changes in plasma proteins in disease cause aggregation of red cells in blood. The aggregates (rouleaux) sediment, before clotting can occur, leaving a clear upper plasma layer. The fibrin clot involves the entire volume of the sample. Subsequently, the clot contracts expressing yellow serum. A clot from a normal person shows two compartments, red (sanguine) and black (melancholic). An extra layer (phlegm), due to the clotted plasma, is seen in the clot of blood from a diseased person. |
In recent centuries the phenomenon of rouleau formation has been studied in glass tubes using blood treated with an anticoagulant. Thus, the sedimentation of the red cells is easily observed. The aggregation is associated with the changes in concentration of various plasma proteins which accompany a variety of diseases. The phenomenon can be induced artificially with blood from a normal subject, either by adding one of a variety of polymers (e.g., polyethylene glycol (PEG), polyvinylpyrollidone, gelatin, dextrans, polylysine, hyaluronic acid, [10, 11, 30, 47]), or by preheating serum to generate albumin polymers, or simply by concentrating serum slightly by ultrafiltration [22]. The disc-shaped red cells are not aggregated by random cross-linking, as in the case of agglutination by bivalent antibodies, but instead form precise "piles of coins" (Fig. 2). |
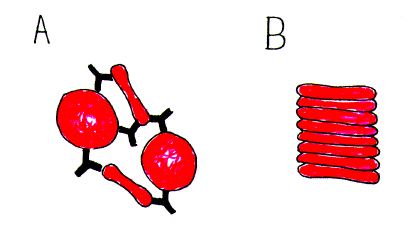
| Fig. 2. Distinction between (A)
aggregation of red blood cells by an extracellular ligand (agglutinating antibody) and (B)
aggregation which does not involve a cross-linking agent.
In (A) the biconcave disk-shaped red cells are cross-linked by Y-shaped bivalent antibodies (not drawn to scale). In (B) the red cells adopt the energetically most favourable pile-of-coins conformation (rouleaux), which sediment rapidly. |
|
The aggregation shows specificity, "like" aggregating with "like" (see later). Thus, it has been concluded that the aggregation is a response of the red cells to some change in the surrounding medium that does not involve cross-linking by an extracellular ligand [20, 30]. Whereas the aggregates with specific antibodies tend to be stable, rouleaux can be disassembled by gentle shaking. Similar weak interactions may be involved in some types of specific aggregation involving nucleated cells [9, 39]. 4. Self-Aggregation of Macromolecules Self-aggregation of macromolecules in solution can be induced by increasing their concentration. Lauffer [ 31, 32] studied the phenomenon using the coat protein of tobacco mosaic virus (TMV). He showed that aggregation involves the liberation of water molecules bound to the macromolecules. Thus, while it might appear, from the observed aggregation, that entropy is decreasing, the increase in disorder of the liberated water molecules more than compensates for the increase in order of the macromolecules. System entropy is increasing.If the aggregation is entropy-driven (endothermic), then it should be promoted by a small increase in temperature. Aggregation can indeed be induced by increasing the temperature over a narrow range, much lower than would be needed to denature the protein (Fig. 3). Similar results were obtained in studies with haemoglobin-S [31]. Both hydrophobic and hydrophilic proteins can be aggregated by small temperature increases [34]. |

|
Fig. 3. Effect of temperature on solutions of TMV coat
protein.
An electron micrograph of a sample (0.1 mg/ml) held at either (A) 4oC or (B) 23oC. (Reprinted from "Entropy-Driven Processes in Biology" by M. A. Lauffer [31].) |
|
5. Specificity of Self-Aggregation In experiments with TMV coat protein, aggregation could be induced by increasing the concentration either of the protein itself, or of one of a variety of other macromolecules in the surrounding medium [7, 33]. Since only the coat protein was present in the aggregates, the aggregation was self-specific. Similar results were obtained in experiments with haemoglobin-S. The surrounding molecules which induced the aggregation of haemoglobin-S were excluded from the aggregates (for references see [37]). In experiments on rouleaux formation, aggregation was also induced by increasing the concentration of macromolecules in the surrounding medium. Since red cells can be easily observed and labelled, it was possible to demonstrate the specificity of the aggregation by light microscopy. In mixtures of red cells from different species, cells of the same species preferentially aggregated with each other (Fig. 4; [20, 47]). Edwards [9] noted that the self-aggregations of molecules and cells share many features suggesting a common mechanism. |

| Fig. 4. Specificity of rouleaux formation. A rouleaugenic
agent is equally active in aggregating red blood cells from animal species A or B.
When the two cell populations are mixed and then treated with the rouleaugenic agent the resulting rouleaux should either contain both A and B red cells (indicating non-specific aggregation), or all A and all B red cells (indicating specific aggregation). The latter alternative is found experimentally. |
|
Addition of increasing quantities of certain "volume excluding agents" (e.g., PEG; [37]) to protein mixtures results in the selective aggregation and precipitation of distinct molecular species at distinct PEG concentrations [29]. It seems probable that there is first a specific like-with-like entropy-driven aggregation, followed by the sedimentation of the aggregates.
6. Proteins Exert a Collective Pressure in the Crowded Cytosol From the above it seems that, if one were to take a mixture of various proteins (A, B, C, D, . . . ) in aqueous solution and subject it to concentration by ultrafiltration, at a certain critical concentration one of the proteins, say A, would self-aggregate. In this process each molecule of A would loose some of its bound water. In the absence of the surrounding proteins (B, C, D, ... ) much higher concentrations of A would be required for aggregation to occur. Thus, a group of proteins collectively exerts a "pressure" (the binding of water) tending to force individual protein species to self-aggregate and give up their bound water. A protein species with the greatest propensity to self-aggregate (a function of factors such as structure, molecular size, and initial concentration), would aggregate first. As the degree of concentration increased, other protein species would follow suite. I have argued that, in disease, just as changes in an extracellular fluid (blood plasma) might create a pressure to drive red blood cells (and perhaps extracellular pathogens) into self-aggregates, so there might also be similar changes in an intracellular fluid creating a pressure to drive intracellular pathogens into aggregates [ 13]. The heat-shock proteins, which in some systems may exceed 10 % of total cell protein (for references see [38]), would play an important role in this. The concept of the "crowded cytosol" implies that much intracellular water is bound to proteins [23], so that there would always be a strong standing pressure to drive into self-aggregation any macromolecular species which exceeded the solubility limits imposed by the macromolecules surrounding it. Each individual macromolecular species would both contribute to the pressure and be acted upon by the pressure. Thus, each macromolecular species would have this collective function as well as a specific function. Both functions could affect phenotype and hence influence selection by evolutionary forces.7. Fine-Tuning of Intracellular Protein Concentrations The concentration of a protein in the cytosol is an important attribute of that protein which is not necessarily related, in any simple way, to the specific role the protein might play in the cell. A protein has evolved to carry out a specific task. On grounds of economy, it might be supposed that evolutionary forces would have pressed for a maximization of specific activity (activity/unit protein). This would minimize the necessary concentration of the protein. However, there is no particular virtue in minimizing the concentration of a cytosolic protein. Provided its concentration is not extreme, a protein itself does not "burden" the cell. Associated with each protein there might be a metabolic burden, which would reflect the energy required to synthesize and degrade the protein. To minimize this burden there might be an evolutionary pressure favouring protein stability, but stability alone does not determine the steady-state concentration of a protein. If the ability to exert a pressure to drive other proteins from solution is an important attribute of a protein (collective function), then the steady-state concentration of the protein might tend towards the maximum compatible with the protein remaining in simple solution without self-aggregation. This would tend to counteract any tendency to maximize specific activity. (However, a high specific activity might be advantageous in creating a "margin of safety" against a decrease in protein concentration; [ 18] (Click Here)Evolutionary forces (mutation, selection, drift) promote the survival and replication of one allele rather than another. Sequence differences between alleles occur not only in protein-encoding regions, but also in other genetic regions which determine factors such as transcription rate and mRNA stability. Differences in protein-encoding regions can affect specific function and/or protein lifespan. Differences affecting steady-state mRNA levels can affect protein synthesis rates. Thus, the steady-state protein concentration would be affected by sequence changes in various parts of the gene. These would often be independent of sequence changes affecting specific function. The net result of evolutionary forces acting on a gene would be a protein of a certain specific activity, which might be less than the maximum possible. Over evolutionary time, the concentration of the protein would have been fine-tuned to the concentrations of the other proteins with which, from generation to generation, it had shared a common cytosol. In this circumstance a new mutation in the protein could result in aggregation (e.g., haemoglobin-S). There are the special cases of proteins whose concentrations vary dramatically with the physiological state of the cell and proteins which form stoichiometric complexes with other proteins. In the case of physiologically regulated proteins, fine-tuning would be with respect to the maximum concentration attained in the cell. In the case of proteins which form a multiprotein complex, the whole complex would act as an evolutionary unit with respect to concentration fine-tuning. 8. Intracellular Self/Not-Self Discrimination Why fine-tune? I have proposed that an important factor favouring the very precise fine-tuning of cytosolic protein concentration would be the need to discriminate intracellularly self-proteins from foreign proteins (such as might be encoded by a virus) [ 13-17]. A system for intracellular self/not-self discrimination could have evolved when the first unicellular organisms arose and were confronted with the first prototypic viruses.Three differences between the genes of a host cell and the genes of a virus could be exploited to discriminate between self and not-self:
In this light it has been proposed that proteins of an intracellular pathogen more readily exceed the solubility limits imposed by the host proteins in the crowded cytosol. Thus, they are preferentially aggregated, self with self [13-17]. The aggregates might then trigger host cell death by some prototypic apoptotic process [50], so that intact virus particles would not be formed or released to affect other cells. These cells might be the "kin" of the initially affected cell, so that the altruistic destruction of host cell genes might further the propagation of copies of the same genes in other cells [25]. Associated with the development of this self/not-self discrimination system would be the development of sets of proteins with the attributes of some modern heat shock proteins [13]. Some of these would act as chaperonins maintaining the structure of self proteins in order to prevent inadvertent aggregation [24]. This would permit fine-tuning of concentration to approach even closer to the threshold (beyond which aggregation would occur). To prevent inadvertent aggregation there would also be a certain margin for error, so that a protein might have to "stick its neck out" to the extent of undergoing a two-fold or greater increase in concentration before aggregating and triggering the prototypic apoptotic process. 9. Heat Shock Proteins Sustain Aggregation Pressure Since the aggregation of not-self molecules would be entropy-driven (endothermic), it would also be produced by a rise in temperature [31]. Thus, the adaptive response of a host to infection by an intracellular parasite might involve locomotion to a region of higher temperature, or a metabolic change to increase temperature endogenously. These responses are documented for various multicellular organisms, including insects [2]. Either as a result of this rise in temperature [5], or by some other mechanism, many organisms show a well developed heat shock response [38]. This involves an elevation in concentration of the heat shock proteins and a concomitant small decrease in the concentration of the majority of self proteins. The latter decrease can be seen as an adaptation to prevent the inadvertent aggregation of self proteins. Depending on the "margin of safety" of specific activity [18], specific function might not be greatly impaired. However, the pressure to drive not-self proteins into aggregates would have decreased. Thus, one role of the heat shock proteins would be to replace the normal spectrum of self-proteins, so sustaining the pressure. The heat shock response can be seen as a manifestation of a sophisticated intracellular host system which evolved, like the extracellular immune system, to combat foreign organisms [6, 35]. It is a rapid response so that there is little time for resistant variants of a rapidly replicating pathogen to arise. However, some pathogens might have evolved mechanisms to prevent aggregation by preventing the induction of heat shock proteins [40]. Others might have evolved mechanisms for interfering with the host-cell response once the self/not-self alarm had rung. One such mechanism might involve inhibiting the expression of MHC proteins [4], whose role is discussed below. An intracellular self/not-self discrimination system which merely trips a prototypic apoptotic system, might have sufficed for unicellular organisms. In a multicellular organism apoptosis of an originally infected cell would prevent the infection of other cells by the progeny of the original virus. However, an organism which could learn from this experience would be better prepared against future viruses than an organism which had no such learning capacity. The involvement of MHC proteins and cytotoxic T cells can be understood in this light. The aggregation of not-self proteins, instead of directly tripping apoptosis, would activate a system leading to the display of peptide fragments at the cell surface in association with MHC proteins. The MHC-peptide complexes would then be recognized by a few T cells whose receptors had a sufficient degree of specificity. The latter cells would then multiply creating a clone. Members of the clone would patrol the body in anticipation of future infections by the same virus. Thus, an immunological memory against the virus would have been created. Pyrexia would have evolved to make it easier to drive a foreign protein out of simple solution as its concentration approaches its individual solubility limit. The diminution of the concentration of normal cytosolic proteins in the course of the heat-shock response would decrease the probability of aggregation of self proteins and hence of the generation of self peptides capable of competing with not-self peptides for binding to MHC proteins. By increasing in concentration at a time when the pressure to aggregate is enhanced, heat-shock proteins might themselves become vulnerable to degradation and might generate peptides which would associate with MHC proteins. |
This possibility, that the intracellular self/not-self discrimination mechanism might be error-prone with respect to certain self proteins, was discussed previously [15]. It is predicted that peptides from these proteins would associate with MHC proteins at the stage of thymic education, thus eliminating the corresponding T cell clones. By lowering the concentrations of most normal proteins, the heat-shock response would shift the population of self proteins at risk from the broad range of normal intracellular proteins to a very narrow range, which would include the heat-shock proteins themselves. This shift in range would minimize the number of "holes" in the T cell repertoire created by deleting self-reacting T cells. Some heat-shock proteins, however, appear able to provoke the production of specific T cell clones [3]. The presence of heat shock proteins at a time when a foreign protein was being aggregated would provide an opportunity for evolution of their involvement in peptide transfer during the processing of fragments of the foreign protein [35]. |
|
11. Other Compartments and Other Interspecies Relationships I have here focused on the cytosol as a major intracellular compartment. However, it is not improbable that similar principles for distinguishing self from not-self apply to other compartments, both intracellular and extracellular [ 21]. I have also focused on the relationship between a host species and a parasite species. However, similar principles would apply to any two species which had not had the opportunity to closely coordinate the concentrations of their gene products over evolutionary time.Some failures to generate hybrids between species might be explained in these terms [19]. The genes of a species undergoing sexual recombination would share a common gene pool and thus would have had the opportunity to fine-tune to each other. The intracellular self/not-self discrimination mechanisms would normally not be tripped in a cross between members of the same species. 12. Further Implications and Experimental Predictions A hypothesis is supported if it leads to predictions which can be verified, or if it provides a unified view which explains experimental findings for which there are currently less satisfactory explanations. Evidence bearing on the existence and effectiveness of an intracellular self/not-self discrimination system might be obtained by comparing the ratios of peptides derived from self and from not-self proteins which could be recovered from the purified MHC molecules of cells infected with various viruses [43]. It would be predicted that virus-derived peptides would be selectively presented. |
Since self/not-self discrimination would occur at the intracellular level, negative selection at the thymus level would be less important. Thus, there should be many peripheral T cells specific for MHC complexes with peptides derived from intracellular self proteins, as noted by Schild and coworkers [46]. |
Another prediction is that it should be difficult to express proteins encoded by expression vectors in a foreign cytosol. This is frequently found. The expressed foreign proteins accumulate as insoluble aggregates. As expected, the aggregates form more readily if the temperature is increased slightly, over a physiological range [44]. Indeed, Nguyen and coworkers [41] showed that expression vector products (e.g., luciferase) lose activity much more when heated to fever-level temperatures in vivo in a foreign cytosol, than when heated independently in vitro (Fig. 5). The loss of activity correlates with protein aggregation, not degradation, and occurs at a lower temperature in a Drosophila cytosol than in a mammalian cytosol. The data imply that the foreign cytosol plays an active and species-specific role in the aggregation. |
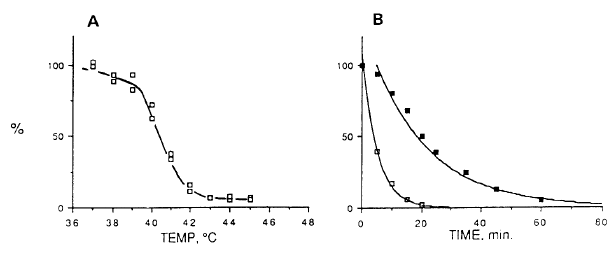
Fig. 5. Effect of temperature on the activity of Photinus pyralis luciferase in cultured rat fibroblasts containing a luciferase-encoding expression vector.
|
|
The fact that some proteins can be expressed successfully in a foreign cytosol, often in cultured eukaryotic cells, would seem to argue against the hypothesis. However, in the absence of cytotoxic T cells there would not necessarily be any negative consequences of the generation of small aggregates. Even if, as a default option, such aggregates could trigger apoptosis, it should be recalled that most culture cell lines and the media they grow in, were developed to optimize cell survival and proliferation, not apoptosis. Transgenic animals containing genes which have not coevolved with host genes, but have followed similar developmental programs, should provide good model systems for studies of the factors affecting intracellular aggregation. Stouthammer and coworkers [48] observed that a variety of
intracellular microbial parasites can be eliminated either by antibiotics or by a small
elevation of temperature. This has been rationalized as demonstrating that some feature of
the metabolism or structure of the parasite is more heat-sensitive than any similar host
feature. This seems unlikely for two reasons:
Second, although the parasite might not have been exposed to the antibiotics throughout
the course of evolution, it might well have been exposed to small temperature elevations.
A parasite with a higher replication rate than its host should readily have evolved an
equal resistance to temperature elevation. A more plausible explanation of the apparent temperature-sensitivity
might be that proposed here. The particulate parasites, acting like extreme
macromolecules, would have been aggregated in the cytosol at an elevated temperature.
Indeed, it seems not improbable that temperature-sensitive mutants in general show
impaired function at the restrictive temperature, not because the protein concerned is
intrinsically heat-sensitive at that temperature, but because of its inability to remain
in simple solution in the crowded cytosol at that temperature. Resistance against the
intracellular parasite Toxoplasma gondii has recently been closely correlated
with the induction of heat-shock proteins [ Another prediction of the hypothesis is that proteins should be under evolutionary constraint not only to retain specific function, but also to retain solubility and lifespan (required for their collective function). Thus, organisms synthesizing proteins with mutations to amino acids adversely affecting the latter properties, would be selected against. Molecular properties such as isoelectric point and molecular weight (which affect aggregatability) would be expected to vary less than predicted on the basis of known amino acid substitution rates. This would be observed experimentally as low inter-species variation in two dimensional gel electrophoretic analyses of proteins [36]. Finally, and quite unexpectedly, the hypothesis provides a possible insight into the evolution of X chromosome dosage compensation, genetic dominance and the lethality of aneuploid states, as discussed elsewhere [16-19; Click here]. That so many disparate phenomena can be understand in the terms of one unifying hypothesis is very encouraging. We seem to have come a long way since the ancient Greeks. |
13. Acknowledgements
This work was supported by the Medical Research Council of Canada and the Leukaemia Research Fund of Toronto.
References
[1] Arber W. and Linn S., DNA modification and restriction, Ann. Rev. Biochem. 38 (1969) pp. 467-500.
[2] Boorstein S. M. and Ewald P. W., Costs and benefits of behavioural fever in Melanoplus sanguinipes infected by Nosema acridophagus. Physiol. Zool. 60 (1987) pp. 586-595.
[3] Born W., Happ M. P., Dallas A., Reardon C., Kubo R., Shinnick T., Brennan P. and O'Brien R. Recognition of heat shock proteins and gamma-delta T cell function. Immunol. Today 11 (1990) pp. 40-43.
[4] Browne H., Smith G., Beck S. and Minson T., A complex between the MHC class I homologue encoded by human cytomegalovirus and Beta
2 microglobulin. Nature, Lond. 347 (1990) pp. 770-772.[5] Cosgrove J. W. and Brown I. R., Heat shock protein in mammalian brain and other organs after a physiologically relevant increase in body temperature induced by D-lysergic acid diethylamide. Proc. Natl. Acad. Sci. USA 80 (1983) pp. 569-573.
[6] DeNagel D. C. and Pierce S. K., Heat shock proteins in immune responses. Crit. Rev. Immunol. 13 (1993) pp. 71-81.
[7] Dudman W. F., Precipitation of tobacco mosaic virus by macromolecules, Nature, Lond. 211 (1966) pp. 1049-1067.
[8] Du Pasquier L., Miggiano V. C., Kobel H. R. and Fischberg M., The genetic control of histocompatability reactions in natural and laboratory-made polyploid individuals of the clawed toad Xenopus. Immunogenetics 5 (1977) pp. 129-141.
[9] Edwards P. A. W., Differential cell adhesion may result from non-specific interactions between cell surface glycoproteins, Nature, Lond. 271 (1978) pp. 248-249.
[10] Fahraeus R., The suspension stability of the blood, Acta Med. Scand. 55 (1921) pp. 1-228.
[11] Fahraeus R., The suspension stability of the blood, Physiol. Rev. 9 (1929) pp. 241-274.
[12] Forsdyke D. R., Further implications of a theory of immunity, J. Theor. Biol. 52 (1975) pp. 187-198.
[13] Forsdyke D. R., Heat shock proteins defend against intracellular pathogens: a non-immunological basis for self/not-self discrimination, J. Theor. Biol. 115 (1985) pp. 471-473.
[14] Forsdyke D. R., Early evolution of MHC polymorphism, J. Theor. Biol. 150 (1991) pp. 451-456.
[15] Forsdyke D. R., Two signal model of self/not-self immune discrimination: an update, J. Theor. Biol. 154 (1992) pp. 109-118.
[16] Forsdyke D. R., Intracellular discrimination between self and not-self: X chromosome inactivation compensates for protein concentration, not function. Proc. Int. Congress Immunol. (Budapest) 8 (1992) pp. 331.
[17] Forsdyke D. R., Relationship of X chromosome dosage compensation to intracellular self/not-self discrimination: a resolution of Muller's paradox? J. Theor. Biol. 167 (1994) pp. 7-12.
[18] Forsdyke D. R., The heat-shock response and the molecular basis of genetic dominance, J. Theor. Biol. 167 (1994) pp. 1-5.
[19] Forsdyke D. R., Fine-tuning of intracellular protein concentrations, a collective protein function involved in aneuploid lethality, sex determination and speciation, J. Theor. Biol. (1995) 172, 335-345.
[20] Forsdyke D. R. and Ford P. M., Segregation into separate rouleaux of erythrocytes from different species. Evidence against the agglomerin hypothesis of rouleaux formation, Biochem. J. 214 (1983) pp. 257-260.
[21] Forsdyke D. R. and Ford P. M., Rouleaux formation as an indication of the phase separating ability of plasma, J. Theor. Biol. 103 (1983) pp. 467-472.
[22] Forsdyke D. R., Palfree R. G. E. and Takeda A. Formation of erythrocyte rouleaux in preheated normal serum: roles of albumin polymers and lysophosphatidylcholine, Can. J. Biochem. 60 (1982) pp. 705-711.
[23] Fulton A. B., How crowded is the cytoplasm? Cell 30 (1982) pp. 345-347.
[24] Gething M. J. and Sambrook J., Protein folding in the cell, Nature, Lond. 355 (1992) pp. 33-45.
[25] Hamilton W. D., The evolution of altruistic behaviour, Am. Nat. 97 (1963) pp. 31-33.
[26] Hedrick S. M., Dawn of the hunt for non-classical MHC function, Cell 70 (1992)pp. 177-180.
[27] Hewson, W., in The Works of William Hewson, ed. by Gulliver G. (The Sydenham Soc., London, 1846) pp. 1-90.
[28] Hitchcock D. I., Proteins and the Donnan equilibrium, Physiol. Rev. 4 (1924) pp. 505-531.
[29] Ingham K. C., Precipitation of proteins with polyethylene glycol, Meth. Enzymol. 182 (1990) pp. 301-306.
[30] Janzen J. and Brooks D. E., Do plasma proteins adsorb to red cells? Clin. Hemorheol. 9 (1989) pp. 695-714.
[31] Lauffer M. A., Entropy-Driven Processes in Biology (Springer-Verlag, New York, 1975).
[32] Lauffer M. A., Motion in Biological Systems (A. R. Liss Inc., New York, 1989).
[33] Lauffer M. A.- and Shelaby R. A., Interaction of tobacco mosaic virus and tobacco mosaic virus protein with bovine serum albumin, Arch. Biochem. Biophys. 178 (1977) pp. 425-434.
[34] Leikin S. and Parsegian V. A., Temperature-induced complementarity as a mechanism for biomolecular assembly, Proteins: Struct. Funct. Genet. 19 (1994) pp. 73-76.
[35] Li Z. and Srivastava P. K., Tumor rejection antigen Gp96/Grp94 is an ATPase: implications for protein folding and antigen presentation, EMBO. J. 12 (1993) pp. 3143-3151.
[36] McConkey E. H., Molecular evolution, intracellular organization, and the quinary structure of proteins. Proc. Natl. Acad. Sci. USA 79 (1982) pp. 3236-3240.
[37] Minton A. P., Excluded volume as a determinant of macromolecular structure and reactivity, Biopolymers 20 (1981) pp. 2093-2120.
[38] Minton K. W., Karmin P., Hahn G. M. and Minton A. P., Nonspecific stabilization of stress-susceptible proteins by stress-resistant proteins: a model for the biological role of heat shock proteins. Proc. Natl. Acad. Sci. USA 79 (1982) pp. 7107-7111.
[39] Morris J. E., Steric exclusion of cells: a mechanism of glycosaminoglycan-induced cell aggregation, Exp. Cell Res. 120 (1979) pp. 141-153.
[40] Nagasawa H., Oka M., Maeda K., Jian-Cuo C., Hisaeda H., Ito Y., Good R. A. and Himeno K., Induction of heat shock proteins closely correlates with protection against Toxoplasma gondii infection. Proc. Natl. Acad. Sci. USA 89 (1992) pp. 3155-3158.
[41] Nguyen V. T., Morange M. and Bensaude O., Protein denaturation during heat shock and related stress. E. coli beta-galactosidase and Photinus pyralis luciferase inactivation in mouse cells, J. Biol. Chem. 264 (1989) pp. 10487-10492.
[42] Ohno S., To be or not to be a responder in T cell responses: ubiquitous oligopeptides in all proteins, Immunogenetics 34 (1991) pp. 215-221.
[43] Rammensee H. G., Falk K. and Rtzschke O., Peptides naturally presented by MHC class I molecules, Ann. Rev. Immunol. 11 (1993) pp. 213-244.
[44] Schein C. H., Production of soluble recombinant proteins in bacteria, Biotechnol. 7 (1989) pp. 1141-1149.
[45] Schwartz R., Acquisition of immunological tolerance, Cell 57 (1989) pp. 1073-1081.
[46] Schild H., Rtzschke 0, Kalbacher H. and Rammensee H. G., Limit of T cell tolerance to self proteins by peptide presentation, Science 247 (1990) pp. 1587-1589.
[47] Sewchand L. S. and Canham P. B., Induced rouleaux formation in interspecies populations of red cells, Can. J. Physiol. Pharmacol. 54 (1976) pp. 437-442.
[48] Stouthammer R. Luck R. F. and Hamilton W. D., Antibiotics cause parthenogenetic Trichogramma to revert to sex. Proc. Natl, Acad. Sci. USA 87 (1990) pp. 2424-2427.
[49] Vidovic D. and Matzinger P., Unresponsiveness to a foreign antigen can be caused by self tolerance, Nature, Lond. 336 (1988) pp. 222-225.
[50] Zychlinsky A., Prevost M. C. and Sansonetti P. J., Shigella flexneri induces apoptosis in infected macrophages, Nature, Lond. 358 (1982) 167-169.
End of paper.
| End Note 2001a: Rajan et al. (2001) Specificity in intracellular protein aggregation and inclusion body formation. Proc. Natl. Acad. Sci. USA 98, 13060-65. The authors use fluorescence resonance transfer to show that "in cells, protein aggregation exhibits exquisite specificity." This dramatically affirms Max Lauffer's studies, and is thus consistent with the hypothesis for intracellular self/not-self discrimination presented on these pages. |
|
End Note
2001b: Intracellular Self/Not-Self Discrimination at
the RNA and DNA levels. Recent studies hint at a process for the recognition of foreign mRNAs, resulting in the formation of double stranded RNAs, which trigger the interferon response and would upregulate MHC expression at the time the foreign peptides would be generated (Cristillo et al., 1998, 2001; Bell & Forsdyke, 1999a,b; Lao & Forsdyke 2000b; Pizzi & Frontali 2001). For intracellular self/not-self discrimination at the DNA level, please see the 1996 paper on the origin of species (Click Here). For more on MHC gene evolution (Click Here). |
End Note 2006: The hypothesis that protein concentrations have evolved to contribute, and to respond, to the aggregation pressure exerted collectively by cytosolic proteins, predicts that excess of one protein will cause differential aggregation of others, thus exerting pleiotropic effects (i.e. changes in functions not necessarily related to that of the original protein). This appears to be supported by Gidalevitz et al. (2006) in a paper entitled "Progressive disruption of cellular protein folding in models of polyglutamine diseases," published in Science 311, 1471-1474. |
|
End
Note May 2008:
Another prediction is
that the likelihood of autoimmune diseases, such as systemic lupus
erythematosus (SLE), would be increased if the setting for aggregation pressure
allowed a higher than normal intracellular protein concentration.. If
it is correct that the adaptive value of sex chromosome dosage
compensation is the control of that setting (Click Here),
it follows that disorders of sex chromosome dosage compensation that
increase the probability of aggregation are more likely to predispose
to the development of autoimmune disease.
Noting the high incidence of SLE in human females (chromosomes XX), Hai-Feng Pan and colleagues have recently drawn attention to observations that the incidence of SLE is higher in XXY males (Klinefelter's syndrome), and low in XO females (Turner's syndrome). They suggest that in normal females there may sometimes be a failure to completely inactivate one of the X chromosomes (to make the dosage of X gene products the same in males and females). In these females a higher than normal aggregation pressure would be expected. "Reactivation of inactive X chromosome: a potential culprit and therapeutic target for systemic lupus erythematosus" by H. F. Pan, D. Q. Ye & X. P. Li. (2008) Medical Hypothesis 70, 1231-1232. "A role for sex chromosome complement in the female bias in autoimmune disease" by Deborah L. Smith-Bouvier et al. (2008) J. Exp. Med. 205, 1099-1108. |
| End Note Nov 2008: Hai-Feng Pan and his colleagues further marshall the evidence for failure to completely inactivate one of the X chromosomes in SLE in a new article (Medical Hypothesis 72, 99-109). The above mentioned study of Smith-Bouvier and her colleagues is described. The fact that failure to inactivate an X chromosome in females would increase the aggregation pressure is now acknowledged: "In these females a higher than normal aggregation pressure would be expected." However, that this enhanced aggregation pressure might influence intracellular self/not-self discrimination is not yet conjectured. |
End Note August 2012 Differential aggregation being the postulated means by which intracellular self/not-self discrimination is triggered, it would be predicted that the primary generation of aggregrates would precede subsequent modifications of the aggregates that are associated with transport to proteosomes, etc. These modifications include ubiquitination and phosphorylation. Recent studies indicate that these occurs after aggregation, suggesting a necessity for prior aggregation so that these downstream events can occur. Li H-Y et al. (2011) Hyperphosphorylation as a defense mechanism to reduce TDP-43 aggregation. PLOS One 6 (8) e23075. Gong B, Kielar C, Morton AJ (2012) Temporal separation of aggregation and ubiquitination during early inclusion body formation in transgenic mice carrying the Huntington's disease mutation. PLOS One 7 (7) e41450. |
|
End Note May 2014:
For growing evidence that intact proteins must be formed first before
their selection for degradation to peptides see EBV page
(Click Here) Aggregation
of these intact proteins is favoured when they are large
and abundant, the latter being likely to correlate with
gene expression level. In an approach that can "discriminate between
sampled and non-sampled proteins," Hoof et al. (2012) provide evidence on
this: "Fortier et al. (2008) observed that MHC-presented peptides are preferentially derived from highly abundant mRNAs. Our analysis confirmed the impact of gene expression reported earlier by Fortier et al., but in addition, our results suggest that protein abundance carries more information for the prediction of protein sampling [for MHC presentation as peptides] than transcript levels do." Hoof, van Baarle, Hildebrand & Kesmir (2012) PLOS Computational Biology 8, e1002517. |
|
End Note
Feb 2018 Peptides from Large and Abundant Proteins
A more extensive mass spectroscopic study of pMHCs adds further support to the view that proteins are not randomly selected for peptide presentation (Pearson et al. 2016). There is "preferential selection of MAPs [MHC class I associated peptides] from a limited repertoire of proteins with distinctive features." The authors emphasize protein size and abundance as key attributes of the donating proteins: "We found that gene expression was the most informative variable, followed by protein length." Here there is emphasis of a distinctiveness of the originating proteins, rather than of the peptides that might derive from these proteins. In other words, the key intracellular self/not-self discrimination event occurs at the level of intact proteins. Pearson et al. (2016) MHC Class I-associated peptides derive from selective regions of the human genome. Journal of Clinical Investigation 126: 4690-4701. |
![]()
Next: For Aggregation-Induced Intracellular Alarm Signals (1999). (Click Here)
For Chaperone Mode-Switches and Double-Stranded RNA (2000). (Click Here)
For Polymorphism and Self/Not-self Discrimination (2001) (Click Here)
For Immunity as a Function of the Unicellular State (2002) (Click Here)
For Heat Shock Response and Self/Not-Self Discrimination (2005) (Click Here)
For X-Chromosome Reactivation and Self/Not-Self Discrimination (2009) (Click Here)
For Acknowledgements (Click Here)
Return to Theoretical Immunology Index (Click Here)
Return to Home-Page (Click Here)
This page was established circa 1999 and was last edited 28 February 2018 by Donald Forsdyke.