|
To really understand evolution we must
first understand the historical development of ideas on evolution. But to really
understand its history, we must first understand evolution.
|
OVERVIEW

Introduction: Species Barriers
Origin of Species (Darwin 1859),
Revisited (Romanes and Gulick)
Two Levels of Information
in DNA (Romanes and Bateson)
Haldane's Rule
Non-Genic ("Chromosomal")
Speciation (Bateson and Goldschmidt)
Grant Allen, George Romanes and
Stephen Jay Gould
Heredity as Information Transfer (Butler)
Darwin's 'Weak Point'
SELECTED PAPERS on the Four Black Boxes: 1. Variation
2. Heredity
3.
Phenotypic ("Natural")
Selection/Isolation
Hybrid Sterility (Darwin 1862)
Variation (Hooker 1862)
Pangenesis (Darwin 1868)
Inutility of Characters
(Gulick 1872)
Natural and Artificial
Selection (Belt 1874)
Inutility
of Characters. Paradox of Sex. Random Drift (Delboeuf 1877)
- 4. Reproductive ("Physiological")
Selection/Isolation
An Unnoticed Factor in
Evolution (Catchpool 1884)
Physiological Selection (Romanes 1886)
Physiological
Selection (Romanes 1887) Embryo
Transfer (Heape 1890)
- "Physiological" Selection/Isolation, the "Chromosomal"
basis
Hybridism and the Germ-Cell
(Guyer
1900, 1902)
Cytological Basis for the
Mendelian Laws (Cannon 1902) 
Chromosomes of the Germ Cells
(Montgomery 1901)
Chromosomes in Heredity (Sutton 1903)
Two Levels of Genetic
Information (Bateson & Saunders 1902)
Heredity and Variation in Modern Lights
(Bateson 1909)
A Phenomenon of Arrangement
(Bateson 1914)
Chromosomes, Polyploidy and Why Sex Evolved
(Winge 1917)
The Blind Watchmaker. Review (1987)
Speciation in Retroviruses (1995)
Origin of Species (1996)
Thinking about Stem-Loops (1998)
Two Levels of Information in DNA (1999)
Haldane's Rule (2000)
Non-Genic (Chromosomal) Speciation (2003)
Allen, Romanes and Gould (2004)
Chromosomal Speciation: A Reply (2004)
Heredity as Information Transfer (2006)
Positive Selection of Synonymous Mutations Initiates Species Divergence (2007)
Darwin's 'Weak Point' (2010)
Acknowledgements
Other Web Sites
Introduction:
Species Barriers
The idea of barriers against members of other species
was evident in the
nineteenth century in the context of infectious disease (Click here). These barriers are both external
(e.g. hygiene), and internal (e.g. the immune response). Internally, our bodies ("self") can detect and destroy members of other species ("not-self"; i.e. viruses, bacteria, protozoa).
Another, no less
subtle form of self/not-self discrimination, involves our detection of a mate ("near-self") who will be our "physiological
complement" such that the union will produce healthy offspring ("hybrids"). An incestuous relationship with a close relative
("too near-self") will probably result in less
healthy offspring. On the other hand, extreme out-breeding, such as with an ape
(not-self), is prohibited by species barriers.
There is more to this
than just the inability to copulate (the gamete transfer barrier). There are both
external components (e.g. mate choice), and
internal components. Even if the male sperm could meet and
fuse with the female ovum, the resulting cell ("zygote")
might be unable to grow from embryo to an adult organism (developmental barrier resulting in "hybrid inviability"). Even if these transfer and developmental
barriers were overcome, in the gonad (testes, ovary) the two sets of parental chromosomes
might be unable to pair for gamete-production (gonadal barrier resulting in "hybrid sterility"). Our modern understanding of these barriers
began with Charles Darwin's The Origin of Species (1859).
The debate on the
primary mechanism of "speciation" continues to this
day. The issues involved are complex.
- What are the relative importances of the three barriers
(transfer, developmental, gonadal) in keeping species separate ("reproductively isolated")?
- Do the barriers appear sequentially, and if so, which
appears first?
- Are there fewer barriers between allied species,
than between distantly related species, or does one barrier just replace another barrier?
- Are the barriers we find between closely related
(allied) species
indicative of the first barriers to appear?
- Does a barrier arise suddenly in an all-or-none fashion,
or does it arise slowly, so that reproductive isolation is initially only partial?
- If one barrier is replaced, does it disappear completely?
- Is there a group of species, members of which, for some
reason related to their biology, have not progressed beyond the first barrier?
- What is the molecular basis of each barrier?
- While we recognize that natural
selection "succeeds" when more fertile offspring are produced,
is there a reproductive barrier that can achieve
this as part of a process that, in extreme form, is independent of
natural selection?
|
Four years after the death
of Darwin in 1882 a major advance was made by Darwin's close research
associate, George John Romanes. Whereas the work of Gregor Mendel (1865) may have been
unrecognized until 1900 because of the relative obscurity of its originator and his
location, George Romanes was at centre stage. |
|
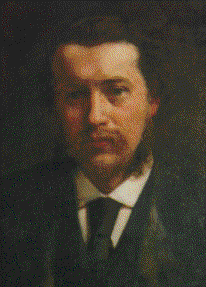
|
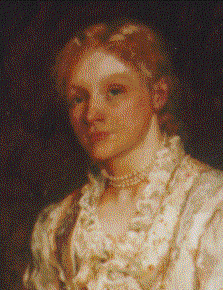
|
|
George Romanes and, his wife and biographer, Ethel
Romanes Photographs of these portraits were kindly provided by their grandson
Giles Romanes (also the grandson of Almroth Wright). The portrait on the
left is signed by T.H. Huxley's son-in-law, John Collier (1850-1934), who, at
Romanes'
suggestion, was commissioned by the Linnean Society to provide the famous 1881 portrait of
Darwin. |
Yet
Romanes'
contributions to evolutionary theory are only just gaining recognition. Perhaps for the
reason given at the top of this page, Biohistorians have not found it easy to tell the
story. John Lesch observed in 1975 that: |
"The development of
evolutionary theory in the two decades from Darwin's death to the turn of the century
remains very largely terra incognita for the historian." |
| William Provine concluded in 1986 that: |
|
"Evolutionary biology in the period 1859-1925 is
extraordinary complex".
|
These web pages
makes available some of the primary documentation in the area, so that you can try
to sort out the issues for yourself. It is obviously biased towards my own
particular evolutionary views, but sites giving alternative perspectives are
easily "googled."
D.
R.
Forsdyke 1998 |
The Origin of Species, Revisited:
A Victorian who Anticipated Modern Developments in Darwin's
Theory
By Donald R. Forsdyke
Queen's
Quarterly (1999) 106,
112-133 (With copyright permission from the Editor, Boris Castel.) 106,
112-133 (With copyright permission from the Editor, Boris Castel.)
In his later years Charles Darwin's closest professional
relationship was with George John Romanes, to whom he entrusted the burden of his life's
work. Four years after Darwin's death, Romanes published a theory of the origin of species
by means of "physiological selection". This
resolved the major problems in Darwin's theory, but replaced them with a "peculiarity" of the reproductive system which would allow
selective fertility between "physiological complements".
To most of his contemporaries, and those who came afterwards, this did not convey much.
However, bioinformatic analysis of DNA sequence data emerging from genome projects now
allows an interpretation. Surprisingly, the words of a "pore
flahr gel" can help us understand work which Alfred Wallace, with the
unknowing aid of a Kingston lady, had condemned to a century of obscurity. |
1848 was a good year for the Reverend George
Romanes, professor of
classics at Queen's College, Kingston, Ontario. He inherited a "considerable
fortune" and his third, and most illustrious son, George John Romanes, was
born. The Presbyterian minister had been in Canada for 14 years, and "relieved of any necessity to continue the duties of his chair",
he returned to Europe (1850), eventually settling in London.
The children, two
surviving boys and two girls, grew up as free spirits. The publication of Charles Darwin's
The Origin of Species by Means of Natural Selection
when George John was eleven, went unnoticed. Some last-minute private tutoring facilitated
George John's entrance to Cambridge University, where he bloomed. His initial interest in
theology, gave way to a life-long interest in biology while maintaining his spiritual
concerns. The biography relates that he "finally abandoned the
idea of a profession" (medicine or the church), and "resolved to devote himself to scientific research."1,2
In 1874 Romanes
published some of his views on evolution in the scientific journal, Nature3. Darwin sent him "a friendly little note" and invited him to visit. |
"From that time began an unbroken friendship, marked on one side by
absolute worship, reverence, and affection, on the other by an almost fatherly kindness
and a wonderful interest in the younger man's work and in his career. ... Mr. Darwin met
him, as he often used to tell, with outstretched hands, a bright smile, and a 'How glad I
am that you are so young!'" |
 |
Darwin had postulated that organisms with variations conferring
some advantage in the "struggle for existence"
would more likely survive and produce offspring. This process of "natural selection" also discriminated against organisms with
disadvantageous variations. The codiscoverer of evolution by means of natural selection,
Alfred Russel Wallace, held rigidly to the theory as originally formulated, while Darwin
was more flexible. Wallace was also a leading advocate of the credibility of supernatural
phenomena, arguing that natural selection was insufficient to account for the evolution of
the human brain (1869)4, and publishing The
Scientific Aspect of the Supernatural (1866)5, and Miracles and Modern Spiritualism (1874)6.
In 1876 Romanes and
his elder brother James were deceived by a "medium"
who claimed to be able to communicate with spirits. Romanes wrote two letters to the
sceptical Darwin expressing an inclination (short-lived) to believe in the phenomena he
had observed. James, who was 14 when the family left Canada, had a friend in Kingston with
an interest in spiritualism and he sent her drafts of the letters. Romanes later (1880)
expressed doubt publicly concerning "the ascertained facts of
clairvoyance and mesmerism" which had been proclaimed in a letter in Nature7.
This brought a first contact with Wallace8. There were two meetings at which
Romanes made no mention of his earlier credulity. |
Meanwhile, Romanes had
formed a close working relationship with the elderly Darwin. Well aware of inconsistencies
in the theory of the origin of species by natural selection, Darwin had made their
resolution his life's main focus. At the time of his friendship with
Romanes, Darwin was
much
concerned with "Pangenesis"
as a possible explanation for the inconsistencies9.
Pangenesis suggested that the testes and ovaries (gonads) were merely collecting centers
for hereditary (and perhaps acquired) information dispersed about the adult body as
independent units which he called "generative elements" or "gemmules".
In an 1875 letter he wrote to Romanes: |

|
"I hope with all my heart that you are getting on pretty well
with your experiments; I have been led to think a good deal on the subject, and I am
convinced of its high importance, though it will take years of hammering before
physiologists will admit that the sexual organs only collect the generative
elements."1, 10 |
| A later letter (1876) began: |
"As you are interested in Pangenesis, and will some day, I
hope, convert an 'airy nothing' into a substantial theory..."10. |
Darwin's correspondence
suggests a sharing, not only of the experimental, but also the theoretical burden of his life's work. Romanes's
experiments to prove the gemmule hypothesis came to nothing, but served to focus his
attention on the gonads.
Following
Darwin's death (1882), Romanes devoted much time trying to find errors in August
Weismann's alternative (but correct) proposal that the germ line (contained in the gonad)
was quite distinct from the rest of the body ("soma").
Romanes's book, An Examination of Weismannism
eventually appeared in 1893, but by this time Weismann's ideas were becoming widely
accepted11. Key features of Weismann's proposal, and the Darwinian alternative,
appear in Figure 1. Here we see that the germ line is part of an unending cycle, and so is
potentially immortal. The soma (that's you and me) lasts for one generation and is then
discarded. |
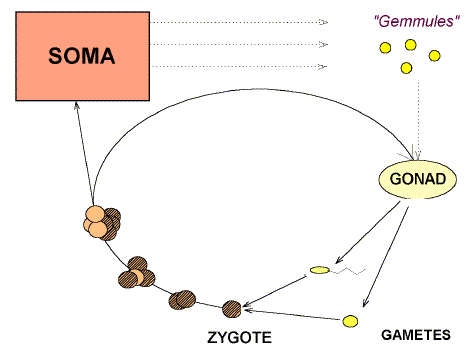 |
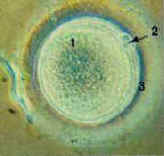
Figure 1. The eternal cycling of
the germ line. Following the gonad-specific process known as "meiosis", the male
gonad (testis) produces male gametes (spermatozoa), and the female gonad (ovary) produces
female gametes (ova). [1 =
ovum. 2 = polar body. 3 = zona pellucida] |
 |
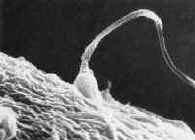
 |
| These unite (fertilization) to produce a unicellular
zygote, which then multiplies and differentiates to produce an adult organism. |
|
|
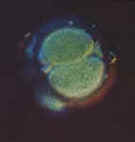
|
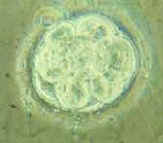
|
Tissues of the organism are either the
"soma" (e. g. liver, brain, kidney), or
the "germ-line" (contained in the gonad). Weismann
proposed (correctly) that the soma (mortal) merely provides a supporting role for
the germ-line (potentially immortal). Darwin proposed (incorrectly) that
throughout life the germ-line would collect "gemmules" of information from the
soma.
The germ line cycle can be interrupted, so that an organism is "reproductively isolated", either because the gametes cannot
meet ("transfer barrier"), or because the zygote does not
develop to produce a healthy adult ("viability barrier"), or
because meiosis fails in the gonad ("sterility barrier").
|
Romanes died in 1894 at age 46, and is remembered today mainly as Darwin's
protege and
for the annual Romanes Lecture series he endowed at Oxford. However, Darwin's mantle
had been safely entrusted. Four years after Darwin's death Romanes published an
extraordinary paper in the Journal of the Linnean Society,
entitled "Physiological Selection: An Additional Suggestion on the Origin of
Species"12. In a formidable display of deductive reasoning, paralleled
only by that of Darwin himself, Romanes resolved the three major problems in Darwin's
theory -- "inutility of characters", "blending inheritance", and "hybrid
sterility". We are concerned here mainly with the latter.
Darwin's
theory was based on observations of plant and animal species "under
domestication". Darwin thought it appropriate to extrapolate
these observations to natural species. Yet he was aware (and the biologist Thomas Huxley
constantly reminded him) that, if crosses between members of closely related natural
species were possible, the "hybrid" offspring were
invariably sterile. If we consider
the horse and the ass as members of separate natural species, when a cross is made between
them the resulting mule, although otherwise healthy, has maldeveloped testes or ovaries,
and cannot reproduce.
Conversely, the
offspring of crosses between members of "species"
created by man (the breeder and horticulturist), are invariably
fertile. If one defines a species as a group of organisms which do
not breed (produce fertile offspring) with members of other species (e.g. cats do not
breed with dogs), then "species" arising under
domestication (e.g. poodle and bulldog) are not true species even though they may differ
greatly from each other anatomically; they are "varieties",
or "races". Hence Darwin confronted a serious
problem.
Returning again to
Figure 1, we see that failure to produce offspring may have three fundamental causes. The
germ-line cycle may be interrupted because:
- (i) The sperm and ovum (gametes) are unable to reach each
other or will not fuse to form the zygote.
- (ii) the zygote cannot develop into an adult organism, so
that the "soma" is not present to support the
survival of the germ-line in the gonad.
- (iii) The formation of gametes in the gonad is impaired.
|
These are sometimes referred to as the transfer barrier, the viability
barrier, and the sterility barrier,
respectively.
Romanes focussed
primarily on the sterility barrier, often the only
barrier separating members of closely related
species (i. e. species likely to have arisen recently from a common originating species).
He suggested that, like cells of all other organs and tissues, germ line cells might
undergo random variations. Variations, for example, in height or eye colour were familiar
to everyone. No one knew what caused variations, but no one doubted their existence.
Romanes
emphasized one possible class of variation affecting germ line cells, which would make an
organism less fertile with other members of the species but not influence somatic
characters. Normally the loss of fertility would be highly disadvantageous because the
organism would leave no offspring. However, he further argued that if two organisms
underwent the same type of variation,
they would still be fertile between themselves.
They would be "physiological complements". Hence,
at any time-point a species would consist of the parental group (comprising the majority
of species members), and numerous small variant groups. Members of each variant group
would be less fertile with members of the parental group and of the other variant groups.
Members of each group, to varying degrees, would be "reproductively
isolated" from members of the other groups, but remain quite fertile with each
other. Hybrid sterility would be an extreme manifestation of the phenomenon.
Romanes demonstrated
that irrespective of any selective
factors in the environment (no "survival of the fittest"
required), members of the reproductively variant groups would tend also to be somatically
variant, just as members of the parental group would tend to be somatically variant (e.g.
changes in height or muscular strength). In the absence of environmental selection,
however, somatic variations of members of the large
parental group would not be sustained, because these members were crossing freely with
each other, a process which tended to blend and neutralize variations. For example,
although humans vary in height, tall people cross with small people and the average height
tends to remain constant.
On the other hand,
variations within a small variant
group would not be subjected to this "swamping"
effect due to blending with members of the parental group (i.e. there would be less
non-variants to dilute the variation). Because of this reproductive isolation, the
variation would be sustained. If, by chance, the variation happened to confer some
advantage to members of the variant group, relative to members of the parental group, that
would be a bonus. The selective advantage conferred would allow the commencement
of classical Darwinian natural selection.
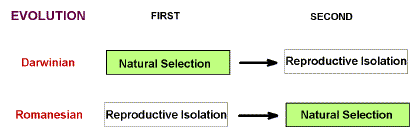
The major difference
between the old Darwinian formulation (natural selection), and the new Romanesian
formulation (physiological selection) was that in the first case, natural selection
preceded reproductive isolation, where in the second case reproductive isolation preceded
natural selection (Figure 2). Reproductive isolation, in any shape or form, would suffice,
but the most usual form of isolation would result from physiological selection. Of course
in a large population it would be very unlikely that a male and a female whose gonads had
undergone the same rare variation would encounter each other. Thus the variation would
probably not be perpetuated. Indeed, as Romanes pointed out, successful speciation was
rare. |
|
|
|
Figure 2. Distinction between the Darwinian and Romanesian Theories of evolution.
|
Wallace criticized the physiological selection theory publicly
(1886)13 and Romanes responded publicly (1887)14. At this time
Wallace, famous as much for his books on spiritualism as for his work on evolution,
visited North America on a lecture tour. In his 1905 biography he relates that he visited
Queen's University, where: |
"After the lecture [on "Darwinism"] some friends of
Principal Grant came in, and ... a lady who was interested in spiritualism ... asked me if
I knew that Romanes was a spiritualist, and had tried to convert Darwin. I told her that I
knew he was interested in ... spiritualism, but that I thought it most improbable that he
had said anything to Darwin. "But," she said, "Professor Romanes's brother
is a great friend of mine, and he gave me the drafts of the letters they jointly wrote to
Darwin. Would you like to see them?" I said I certainly should, and she promised to
bring them the next morning. ... She said I might take notes on the contents as they were
given to her without any restriction."8 |
The
name of the lady is not disclosed, but we know that Wallace had made the acquaintance in
Washington of the family of the Reverend J. A. Allen, whom he later visited at his home in
Kingston. Wallace also took tea at Sir Richard Cartwright's "fine
country house in a spacious park a few miles in the country". |
Wallace was everything that Romanes was not. Wallace was
relatively poor, a socialist (he published Land Nationalization in 188215), an
anti-vaccinationist (he published Vaccination a Delusion
in 189816), an advocate of spiritualism, and, with the death of Darwin, the
preeminent authority on evolution. At the close of the nineteenth century there was a
sense of complacency both in the biological and the physical sciences. It seemed that the
major issues had been resolved, and now it was just a question of sorting out the details17,18.
In this environment authorities carried much weight. published Land Nationalization in 188215), an
anti-vaccinationist (he published Vaccination a Delusion
in 189816), an advocate of spiritualism, and, with the death of Darwin, the
preeminent authority on evolution. At the close of the nineteenth century there was a
sense of complacency both in the biological and the physical sciences. It seemed that the
major issues had been resolved, and now it was just a question of sorting out the details17,18.
In this environment authorities carried much weight.
In a book entitled Darwinism (1889), Wallace modified views previously
expressed in correspondence with Darwin (1868)10, 19. The correspondence
contained a nineteen-point "proof" concerning the
role of natural selection in hybrid sterility, which began: |
"Let there be a species which has varied [note: past tense] into two forms
each adapted to certain existing conditions better than the parent form, which they soon
supplant [Wallace's italics]"10. |
"I demur
to probability and almost to possibility of ... [point 1] as you start
with two forms which are not mutually sterile, and which yet have supplanted the
parent-form". |
| Two decades later in his 1889 book Wallace
admitted: |
"The preceding argument ... [now decreased to eleven points]
depends entirely upon the assumption that some amount of
infertility characterizes the distinct varieties which are in process of differentiation
into species; and it may be objected that of such infertility there is no proof"19. |
Under Wallace's scheme, the event which concerned Romanes -- the initiation
of the speciation process -- already
had happened. Wallace dealt with events subsequent to the process of reproductive
isolation (i.e. maintenance rather than initiation; Figure 3). The idea that the
infertility he noted might relate to what Romanes proposed did not occur to Wallace. In a
separate section of his book he described physiological selection as "another form of infertility," which he then proceeded to
attack19.
Romanes responded
publicly once more in 1890, this time not only with scientific arguments, but also noting
that one should not rely on the judgement of a person of "incapacity
and absurdity" whose past views were highly questionable, namely, on
spiritualism, socialism and vaccination20. Wallace wrote Romanes some sour
private letters, which were later published8. These letters protested the
"appeal to popular scientific prejudice," revealed
Wallace's knowledge of Romanes's earlier flirtation with spiritualism as gleaned from the
Kingston letters, and threatened to make "known the fact of the
existence of these letters and their general tenor." This would show that
Romanes's private judgements did not accord with his public posture of scepticism with
respect to spiritualism. In short, Romanes was a hypocrite. A century later, of course,
there are many reasons to conclude that Romanes's mature
judgements with respect to spiritualism, socialism and vaccination were correct. Perhaps
his judgement on physiological selection was also correct. |
With Romanes's untimely death in 1894, the physiological selection
theory lost its most powerful advocate. The most able of his English contemporaries,
William Bateson
(1861-1926), was deeply involved in detailed study of variations, work given great impetus
by the discovery in 1900 of a 35-year-old paper by Gregor Mendel21. This
forgotten paper provided a foundation for the new science of genetics. In a 1904 address
Bateson extolled the virtues of the "practical man" who will
"stoop to examine Nature"
in "the seed bed and the poultry yard". Bateson seemed not to think
highly of those (unnamed) with a philosophical bent of mind, who were interested in hybrid
sterility achieved by some imaginary form of selection: |
 |
"For the concrete in evolution we are offered the abstract.
Our philosophers debate with great fluency whether between imaginary races sterility grew
up by an imaginary Selection ... and for many whose minds are attracted by the abstract
problem of inter-racial sterility there are few who can name for certain ten cases in
which it has already been observed"22. |
Bateson sent a copy of
his data-laden book, Materials for the Study of Variation23,
to Huxley, who replied (1894):, |
"How
glad I am to see ... that we are getting back from the region of speculation into that of
fact again. There have been threatenings of late that the field of battle of Evolution was
being transferred to Nephelococcygia [meaning nonsense].24" |
 |
In private
correspondence in 1888, Huxley had expressed the view that "even
people like Romanes" did not really understand Darwin's theory, which was why
Romanes had got it "so hopelessly wrong"24.
Thus, Romanes did not have the support of some of the most influential people of the time,
-- Wallace, Huxley, and Bateson. |
While Romanes pursued his evolutionary studies, Conan Doyle was
creating the great detective Sherlock Holmes, who advised: |
"When you
have eliminated the impossible, then whatever remains, however improbable, must be the
truth".25 |
In Holmesian style Romanes had eliminated the "impossible", but
"what remained" seemed so "improbable" that few
could be convinced that physiological selection "must be the truth".
Romanes could not go into specifics as to the cause of the reproductive selection he was
postulating, beyond what he termed a "physiological peculiarity" of the
reproductive system, the most obvious manifestation of which was the phenomenon of hybrid
sterility. Nor could he elaborate upon occasional remarks that speciation (selective
fertility) required the "suitable
mating of 'physiological complements'." To
some, there may not have appeared much difference between this view and the notion of
divine creation. |
Indeed, the outspoken purpose of people such as the botanist
Alexis Jordan, whose observations Romanes used to buttress his case, was to disprove
Darwin in favour of divine creation26. It may not have added to his scientific
credibility that Romanes was the son of a Presbyterian minister, that his major ally was
the Reverend John Gulick,27 and that Romanes, known among his friends as "the philosopher," had written books with such
titles as A Candid Examination of Theism
(1878)28, Mind and Motion
(1886)29, and Thoughts on Religion
(1895)30. In 1879 he had declined an invitation from Huxley, well known for his
agnostic views, to join the Association of Liberal Thinkers24.
I n
1886 Romanes had regarded his studies as merely paving the way for future investigations: |
 |
"My suggested explanation of the origin
of species opens up another and more ultimate problem, namely, granted that species have
originated in the way supposed, what have been the causes of the
particular kind of variation in the reproductive system which the theory requires?"12 |
To
give his Victorian audience a concrete example of one
form of physiological selection, Romanes had mentioned a change in the time of flowering
of a plant, which would restrict the plant to fertilization only by other members of the
species which had undergone the same
change in flowering time: |
"Suppose the variation in the reproductive system is such that
the season of flowering or of pairing becomes either advanced or retarded. ...some
individuals living on the same geographical area as the rest of their species, have varied
in their reproductive system, so that they can only propagate with each other. They are
thus perfectly fertile inter se, while absolutely sterile with all the other
members of their species. This particular variation being communicated by inheritance to
their progeny, there would soon arise on the same area, ... two varieties of the same
species, each perfectly fertile within its own limits, while absolutely sterile with one
another. That is to say, there has arisen between these two varieties a barrier of
intercrossing which is quite as effectual as a thousand miles of ocean; the only
difference is that the barrier, instead of being geographical, is physiological".12 |
Today we would regard such cases as resulting from primary variations in specific genes
which affect the time of flowering. However, Romanes sensed that something more than
specific genes was involved in the general
case of formation of new species by physiological
selection.12
The
Romanes-Gulick correspondence indicates that although acknowledging its relevance, John
Gulick (1832-1923) had not really appreciated the importance of hybrid sterility31.
Thus, his son and biographer, the biochemist Addison Gulick noted in 1932: |
"[John] Gulick's writings of 1887 and thereafter give an
elaborate discussion of different aspects of sterility, though never quite placing mutual
sterility in relief, the way Romanes did, as likely to be the fuse that ignites the whole
powder train".27,
31 |
Romanes
and Gulick had been separately climbing towards the peak of a high mountain, for much of
the time their heads being lost in the clouds. Every-so-often the clouds would have
cleared and they would have been privileged to views, lonely views, which they could
partially communicate to each other, but not to their contemporaries on the slopes below.
In 1890 Romanes wrote to Gulick: |
"It appears to be desirable that, as you and I are the
only two human beings who recognize the full importance of "segregation"
[reproductive isolation] in all its forms, we should submit to each other our views before
publication, in order that we may speak as far as possible with a common voice".31 |
Meanwhile, Bateson was much occupied with overcoming the
opposition to Mendel, and seemed to have forgotten Romanes's work. However, late in
Bateson's career the overwhelming importance of the phenomenon of hybrid sterility dawned
on him, and he began to consider its possible causes. In 1913 Bateson dismissed sterility
associated with certain anther variants in plants as being due to a "simple recessive character," implying that sterility due to
defects in one or a few defined characters was unlikely to be of general significance for the evolution of species.32
Microscopical studies of the gonads of sterile hybrids supported this view.
Normally, the set of
chromosomes carrying genes contributed by the mother pair, on a one-to-one basis, with the
complementary set of chromosomes carrying the genes contributed by the father. Close
identity is essential for this pairing, which is required so that subsequent cell division
and multiplication can proceed, resulting in the formation of gametes (sperms and ova). In
hybrid sterility the pairing is defective and a degenerate gonad results. Thus the
question of the origin of species becomes a question of the origin of hybrid sterility,
which, in turn, leads to the question of the nature of the chromosomal incompatibilities which prevent pairing, or
conversely, the nature of the chromosomal compatibilities
which permit pairing. Reminiscent of Romanes's position, Bateson interpreted hybrid
sterility as reflecting complementary chromosomal "factors": |
"Though we cannot strictly define species, they
yet have properties which varieties have not, and ... the distinction is not merely a
matter of degree. The first step is to discover the nature of the [chromosomal] factors
which by their complementary action inhibit the critical divisions
and so cause the sterility of the hybrid".32 |
Romanes
and Bateson used the word "complementary" in
converse contexts. Romanes's "physiological complements"
were organisms with reproductive systems which were compatible,
so that offspring would be produced. Bateson's "complementary
factors" were components of the reproductive systems which were incompatible, thus preventing the production of
offspring. As we shall see, these were but two sides of the same coin.
That hybrid sterility
was strictly the result of a failure of the pairing of complementary chromosomes was
brought home forcefully by experiments showing that hybrid sterility could be "cured" in certain organisms by artificially doubling the
number of chromosomes. This was because each chromosome now had a partner with which it
could pair (i.e. chromosomes from the mother could pair with each other, and chromosomes
from the father could pair with each other). If the sterility had been due to a defect in
particular genes, this "cure" would be unlikely33.
Romanes's
ally, John Gulick, remained uninfluential despite a major monograph in 190534.
He generated a complex classification of types of sterility, which, combined with his
cumbrous prose, no doubt confused more than it clarified. But Addison Gulick continued the
crusade. In his 1932 biography of his father he wrote: |
"The problem of physiological isolation has
received ... little attention during the last two decades, but the modern picture of the
mechanism of reproduction and the physico-chemical processes that it involves would have a
very large influence on any consideration of the subject today. The immediate cause of
infertility between species would be sought at the present time in either chromosomal
incompatibilities or maladjustments of a serological nature .... From several viewpoints
the old proposition of Romanes seems today exceedingly plausible, that a rather trivial
mutation or group of mutations might set up a barrier of sterility within a species, and
cause the two portions to diverge thereafter into well-marked new
species".31 |
Unfortunately, Addison Gulick's campaign came too late. By 1932 the high ground of
evolution research had been seized by individuals who approached evolutionary questions
from a different perspective. Nevertheless, he attempted to open the argument again in
1938: |
"With the progress of years we find a striking
reinforcement of the scientific cogency of the theory which Romanes and J. T. Gulick
championed; namely that a physiological barrier between two otherwise hardly
distinguishable stocks may occur frequently, and must have the effect of initiating
a train of divergent evolution".35 |
Addison Gulick's efforts were to no avail. Those responsible for
what subsequently became known as "the modern
evolutionary synthesis" seem to have been unaware of the
physiological selection theory. They spent much time juggling "selection
coefficients" and the extent and timing of geographical migrations in order to
provide a mathematical underpinning for evolution by natural selection. Their attempts to
reconcile Darwin's ideas with Mendelian genetics were described in The
Origins of Theoretical Population Genetics by historian William
Provine (1971), who concluded: |
"Thus, with the gap between theoretical models and
available observational data so large, population genetics began and continues with a
theoretical structure containing obvious internal inconsistencies".36 |
Romanes himself had
doubts concerning "numerical computation involving the doctrine
of chances", remarking: |
"In reference to biological problems of the kind now
before us, I do not myself attach much importance to a merely mathematical analysis. The
conditions which such problems involve are so varied and complex, that it is impossible to
be sure about the validity of the data upon which a mathematical analysis is
founded".37 |
This foreshadowed later
attacks on the "arid calculations of the mathematical
population geneticists", which Provine relates.36 The
arch-population geneticist John Maynard Smith in his book Evolutionary
Genetics warned in 1989 that: "To
paraphrase Mr. Truman, if you can't stand algebra, keep out of evolutionary biology"!38
In a letter to Darwin a century earlier, Romanes noted: |
"The mathematicians must be a singularly happy
race, seeing that they alone of men are competent to think about the facts of the cosmos.
... Mathematics are ... the sciences of number and measurement, and as such, one is at a
loss to perceive why they should be so essentially necessary to enable a man to think
fairly and well upon other subjects. But it is, as you once said, that when a man is to be
killed by the sword mathematical, he must not have the satisfaction of even knowing how he
is killed".1 |
In human
societies priestly groups always have tended to seize the high ground. Those who occupy
such ground in modern times not only find it easier to continue their dominance, but come
to act as "gate-keepers", the
peer-reviewers who
decide what the rest will be allowed to read and whose research will be funded. This has
its dangers.
In 1976 Richard Dawkin's The Selfish Gene, which magnificently synthesized
the work of William Hamilton and George Williams, added a fresh perspective to Darwinian
ideas.39-41 However, the new viewpoint was largely biological, and many
inconsistencies remained. Then in the 1990s came the genome project and a deluge of DNA
sequence data from biochemical laboratories. The first results of the bioinformatic
analysis of these data have suddenly given the story fresh impetus. It appears that the
chromosomal incompatibility which allows species to originate either arises from "macromutations" which affect chromosome segments31, 33,
or is the cumulative result of
numerous "micromutations," as proposed by the
present author in 199642. In either case, Romanes was right as he did not
specify the nature of the "peculiarity of the reproductive
system" which would lead to reproductive isolation.
The strength of the
micromutation proposal is that it allows fine degrees of compatibility, so that a rare
reproductive variant would be more likely to find a partially matching physiological
complement. The proposal arose from the perception that DNA, the molecule in chromosomes
which transmits hereditary information, has two levels
of information. There is nothing strange about this. A radio operator has to decide what
message to transmit, and on what wavelength to transmit it. Thus the radio signal contains
primary information (the message) and secondary information (the wavelength). The message
is basically the same whatever wavelength is employed. The role of the secondary
information is to allow discrimination between messages; it allows the receiver to tune
out unwanted messages.
Shaw's 1913 play Pygmalion provides another helpful metaphor43.
If asked to recite a nursery rhyme, the "toff"
Freddy Eynsford Hill, of some social standing, would have said: |
|
"Mary had a little lamb, its
fleece was white as snow...".
|
 |
| However, the flower seller, Eliza
Doolittle, would have said: |
|
"Miree ader li-awl laimb,
sfloyce wors woyt ers snaa ...".
|
Both sentences convey
the primary information that Mary
possessed an immature, white, sheep. In class-conscious England at the beginning of the
20th century, however, the sentences would also have communicated the secondary information that Eliza was of "lower class" cockney origin, whereas Freddy was of "middle class" origin.
We may regard Eliza's
dropped Hs as micromutations. The secondary information, because of such
micromutations, constitutes a "reproductive barrier".
Cockneys tend to marry cockneys and perpetuate cockney secondary information. The middle
class tend to marry into the middle class and perpetuate middle class secondary
information. Thus, Eliza appeared to be reproductively isolated from Freddy largely
because of her language. Although, perhaps with some difficulty, both could understand
each other's primary information, the secondary information (Eliza's dropped Hs)
seemed to constitute a barrier.
Shaw tells us that
Professor Higgins performed the appropriate experiment. He demonstrated that if the
linguistic barrier were removed, then the reproductive barrier would also be removed.
Eliza and Freddy lived happily-ever-after. We can say that they then possessed "linguistic complements", by which we mean that they approached
linguistic identity. Alternatively, we can say that until Professor Higgins had performed
his magic, their reciprocal differences, or "complementary
factors" (Eliza's dropped Hs, and Freddy's included Hs)
kept them reproductively isolated.
It is important to
note that Eliza's cockney accent was a global characteristic of her speech. Each sentence
she spoke was influenced by it. While each sentence provided a discrete
piece of "primary" information, which was localized
in that sentence, the "secondary" information (her accent) was diffused across all
sentences.
We should also note
that once the marriage knot had been tied (i.e. the barrier to reproduction formally
removed), a good chance existed that, even if her accent subsequently regressed (diverged
from Freddy's, so that linguistic complementarity was lost), the marriage (reproduction)
would continue. Thus the initial
secondary information (a reproductive barrier) needed only to be removed temporarily until a more substantial (legal) way of
removing the barrier had been substituted.
This latter point
proves of considerable importance when we move back to the problem of gaining evidence for
two levels of information in DNA. Whatever the initial
isolation process which led to the divergence between ancestral lines leading to, say, the
modern elephant and mouse, it is clear that there is now a more substantial barrier to
reproduction between the two (a gamete transfer barrier). It would probably be fruitless
to examine these species for molecular clues as to the way the initial divergence took
place (Figure 3). In virus species, however, signs of the initial isolation process might
still exist. This, indeed, seems to be the case with viruses with the potential to coexist
in the same host cell42. |
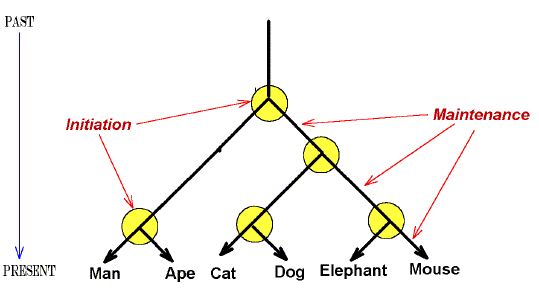
Figure 3. A hypothetical evolutionary tree to show that the initial
evolutionary events required for divergence from prototypic ancestral forms (circled
regions), are distinct from evolutionary events required for the maintenance and further
development of a species. |
The first
critical evidence emerged from studies of insect viruses by biochemist Gerard Wyatt in
1952, and there is now supporting data from analyses of retroviruses and herpes viruses44-46.
We also know that DNA has at least two levels of information. The primary information
occurs in localized "sentences", which are the
genes encoding the various characteristics of an organism. The secondary information is
the molecular basis of Romanes' "physiological peculiarity"
of the reproductive system, and must match between two individuals ("physiological complements") in order that they may reproduce.
A divergence in this secondary information, through accumulation of
micromutations,
appears likely to initiate the speciation process42,47. We should note that
Bateson's careful use of the general term "factors"
(not "genes") allowed him to encompass any form of
information (primary or secondary) which was contained in the chromosomes48.
At
the conclusion of the 1932 biography of his father, Addison Gulick wrote: |
|
"We may anticipate that
the evolutionary problems which were outlined by Romanes and Gulick will again be
reinstated in more perfect form than is today attainable. Until that time arrives it will
remain impossible to estimate the relative importance of the several forces that cause the
progressive transformation of species living under the conditions that nature
provides".31 |
| It has
been a long wait. 35 years passed before the work of Gregor Mendel21 was
recognized in 1900. A century has passed since the posthumous publication in 1897 of
Romanes' masterpiece, the third volume of his Darwin, and
after Darwin, entitled Isolation and
Physiological Selection37. Noting the leisurely pace of
nineteenth century, compared with twentieth century, science, it can be seen that Romanes
was not merely years ahead of his
contemporaries, he was light years
ahead.
However, there are
signs of change. Harvard evolutionist Stephen Jay Gould in a 1980 paper "Is a new and general theory of evolution emerging?" notes a
new readiness to entertain "chromosomal alterations as
isolating mechanisms", and points out49 that: |
"Some of
the new models ... regard reproductive isolation as potentially
primary and non-adaptive rather than secondary and adaptive ... . In ... chromosomal
speciation, reproductive isolation comes first and cannot be
considered an adaptation at all. It is a ... [random] event that establishes a species by
the technical definition of reproductive isolation. To be sure, the later success of this
species in competition may depend on its subsequent acquisition of adaptations; but the
origin itself may be non-adaptive. We can, in fact, reverse
the conventional view and argue that speciation, by forming new entities ... [randomly],
provides raw material for selection"., |
| Of course, Romanes and Gulick would
have warmly agreed with this12,13, 50,51. |
Notes
[For space reasons the notes had to
be abbreviated in the paper published in Queen's Quarterly.]
1 E. Romanes, The Life
and Letters of George John Romanes (London: Longmans, Green & Co.,
1896).
2 C. Darwin, The
Origin of Species by Natural Selection or the Preservation of Favoured Races in the
Struggle for Life (London: John Murray, 1859).
3 G. J. Romanes, "Disuse as
a reducing cause in species", Nature,
10 (1874) 164.
|
Romanes
became a close friend of Norman Lockyer, the
founding editor of Nature,
and had him as a summer guest in Scotland in the early
1880s.
DRF
Aug 2003 |
4 A. R. Wallace, Review of Principles
of Geology by C. Lyell, Quarterly
Review 126 (1869) 187-205.
5 A. R. Wallace, The
Scientific Aspect of the Supernatural(London: F. Farrah, 1866).
6 A. R. Wallace, On
Miracles and Modern Spiritualism (London: James Burns, 1874).
7 G. J. Romanes, ""A
Speculation Regarding the Senses"", Nature
21 (1880) 348.
8 A. R. Wallace, My
Life (London: Chapman
& Hall, 1905), Chapter 36.
9 C. Darwin, The
Variation of Animals and Plants under Domestication. Volume II, 2nd
Edition (London: John Murray, 1875), Chapter 27. [How did Darwin come
upon the idea of pangenesis? For more on this (Click Here).]
10 F. Darwin and A. C. Seward, More
Letters of Charles Darwin (London: John Murray. 1903). [ Haeckel had referred to pangenesis as an "airy
nothing." In similar vein,
Shakespeare in Midsummer Night's Dream
wrote:
"And as imagination bodies forth,
The forms of things unknown, the poet's pen
Turns them into shapes, and gives to airy
nothing
A local habitation and a name." |
11 G. J. Romanes, An
Examination of Weismannism (Chicago: Open Court Pub. Co., 1893).
| It seems
that Weismann eventually came round to adopting something like
Romanes' physiological selection in his theory of "germinal
selection." Thus he wrote (1896): "The variations
presented to personal [natural]
selection must have themselves been produced by the
principle of survival of the fit! And this is effected ...
through such profound processes of selection in the interior of
the germ plasm as I have endeavoured to sketch ... under the
title of germinal selection." |
12 G. J. Romanes, "Physiological
Selection: An Additional Suggestion on the Origin of Species", J. Linn. Soc.(Zool.) 19 (1886)
337-411. [Reproductive isolation due to differences in the time of flowering (plants), or
pairing (animals) he regarded as "extrinsic".
The most important form of reproductive isolation with respect to the origin of species
was considered due to a "peculiarity" of the reproductive system, which he
regarded as "intrinsic".
(For full text -
Click Here)]
13 A. R. Wallace, "Romanes versus
Darwin", Fortnightly Review 46
(1886) 300-316.
14 G. J. Romanes, "Physiological
Selection", Nineteenth Century
21 (1887) 59-80. For full text - Click Here)
15 A. R. Wallace, Land
Nationalization its Necessity and its Aims (London: Trubner & Co.,
1882).
16 A. R. Wallace, Vaccination
a Delusion (London: Swan, Sonnenshein & Co., 1898).
17 L. Badash, "The
complacency of nineteenth century science", Isis
63 (1972) 48-58.
18 A. M. Silverstein, A
History of Immunology (San Diego: Academic Press, 1989), pp. 142-145.
19 A. R. Wallace, Darwinism
(London: Macmillan & Co., 1889), Chapter 7.
20 G. J. Romanes, "Wallace
on Physiological Selection", The Monist
1 (1890) 1-20.
21 G. Mendel, "Versuche uber
Pflanzen Hybriden", Verh. naturf. Ver. in
Brunn 4 (1865) 3-47.
22 W. Bateson, Presidential Address to the Zoological Society; British Association
1904, in William Bateson,
F.R.S. Naturalist. His Essays and Addresses, ed. B. Bateson
(Cambridge: Cambridge University Press, 1928), pp. 233-259.
23 W. Bateson, Materials
for the Study of Variation (London: Macmillan & Co., 1894).
24 T. H. Huxley, Letters, in Life
and Letters of Thomas Henry Huxley, ed. L. Huxley (London: Macmillan
and Co., 1900).
25 A. Conan Doyle, "The Sign
of Four", Lippincott's Monthly Magazine,
(Philadelphia) 266 (1890) pp. 172.
26 A. Jordan, "Remarques sur
le fait d'existence en societe a l'etat sauvage des especes vegetale affine", Congres de l'Association Francaise pour l'Avancement des Sciences,
1873.
27 J. T. Gulick, "Divergent
evolution through cumulative selection", J.
Linn. Soc. (Zool.) 20 (1887) 189-274.
28 G. J. Romanes, A
Candid Examination of Theism (London: Truebner & Co., 1878)
29 G. J. Romanes, Mind
and Motion; and Monism (London: Longmans, Green & Co., 1886).
30 G. J. Romanes, Thoughts
on Religion (Chicago: Open Court Pub. Co., 1895).
31 A. Gulick, John
Thomas Gulick: Evolutionist and Missionary (Chicago: University of
Chicago Press, 1932).
32 W. Bateson, Problems
of Genetics (New Haven: Yale University Press, 1913), pp. 238- 242.
33 T. Dobzhansky, Genetics
and the Origin of Species (New York: Columbia University Press, 1937),
Chapters 7, 9.
34 J. T. Gulick. Evolution:
Racial and Habitudinal (Washington: Carnegie Institute publication
number 25, 1905).
35 A. Gulick, "What are
genes? 1. The Genetic and Evolutionary Picture", Quart.
Rev. Biol. 13 (1938) 1-18.
36 W. B. Provine, The
Origins of Theoretical Population Genetics (Chicago: University of
Chicago Press, 1971).
37 G. J. Romanes, Darwin,
and After Darwin. Isolation and Physiological Selection (London:
Longmans, Green, & Co., 1897).
38 J. Maynard Smith, Evolutionary
Genetics (Oxford: Oxford University Press, 1989).
39 R. Dawkins, The
Selfish Gene (New York: Oxford University Press, 1976).
40 W. D. Hamilton, "The
Genetic Evolution of Social Behaviour", Journal
of Theoretical Biology 17 (1964) 1-54.
[For more on WDH (Click Here)]
41 G. C. Williams, Adaptation
and Natural Selection (Princeton: Princeton University Press, 1966).
42 D. R. Forsdyke, "Different
biological species 'broadcast' their DNAs at different (G+C)% "wavelengths"",
Journal of Theoretical Biology 178
(1996) 405-417.(For full text -
Click Here)
"Micromutations"
were also suggested by Richard Goldschmidt in his Material
Basis of Evolution (1940).
DRF Aug 2003 |
43. G. B. Shaw, Bernard
Shaw. Complete Plays with Prefaces. Volume I. (New York: Dodd, Mead
& Co., 1963). [A character in another Shaw play
"The Doctor's Dilemma" is inspired by Sir Almroth Wright, who
shares with George J. Romanes the honour of being grandfather to Giles J.
Romanes.]
44 G. R. Wyatt, "The nucleic
acids of some insect viruses", Journal of General Physiology 36
(1952) 201- 289.
45 E. C. Bronson and J. N. Anderson, "Nucleotide composition as a driving force in the evolution of retroviruses",
Journal of
Molecular Evolution 38 (1994) 506-532.
46 G. A. Schachtel, P. Bucher, E. S. Mocarski, B. E. Blaisdell, and S. Karlin, "Evidence for selective evolution in codon
usage in conserved amino acid segments of human alphaherpesvirus proteins", Journal of Molecular Evolution 33
(1991) 483-494.
47 More scientific background is provided by references in my 1996 paper (note 42).
See also D. R. Forsdyke, "An alternative way of thinking about
stem-loops in DNA", Journal of Theoretical Biology 192 (1998)
489-504. (For full text
- Click Here) The latter paper provides evidence that pairing of chromosomes
should be influenced by small fluctuations in the species-dependent component of the base
composition of DNA [(C+G)%].
48 H. A. Orr, "Dobzhansky,
Bateson, and the genetics of speciation", Genetics
144 (1996) 1331-1335. I further analyze Bateson's contribution in Journal of Theoretical Biology (1999) 201,
47-61. "Two levels of information in DNA. Relationship of
Romanes' "intrinsic" variability of the reproductive system, and Bateson's
"residue," to the species-dependent component of the base composition, (C+G)%".
[For full text - Click
Here)]
49 S. J. Gould, 1980. "Is a
new and general theory of evolution emerging?" Paleobiology
6 (1980) 119-130.
Lest this quotation be deemed prescient, we
should note recantation by SJG in 2002 (The
Structure of Evolutionary Theory. p. 1003). Here he
stated:
"I do not, in fact and
retrospect ... regard this 1980 paper as among the ... most
cogent ... that I have ever written. ... I then read the literature on speciation as beginning to favor sympatric
[same
country]
alternatives
to allopatric [different
country]
orthodoxies ... and predicted that views on this subject would change substantially,
... . I now believe that I was wrong in this prediction."
DRF
April 2002
There are
uncanny similarities between Romanes and Gould. Both were center
stage, both were well versed in biohistory, both wrote for popular
and scientific audiences, both were attacked by the establishments
of their days, and both contracted cancer in their forties. Modern
chemotherapies may have given Gould another 20 years, whereas
Romanes died at 46.
DRF Aug
2003
|
50 There is a somewhat surrealistic personal twist to this story:
| My bioinformatic work (note 42)
led to a relatively simple view of the origin of species, which made me wonder if
one of the perspicacious Victorians close to Darwin might have
anticipated me.
After following several false trails in my search for a Victorian (e.g.
note 36), I came across Romanes in the fall of 1997 and learned of his Canadian origin.
Now, it so happens that, with my family, I had moved in 1982 to a grey stone house which
forms part of a small block in William Street, Kingston. Shortly after the move we found
that in the 1840s the block had housed Queen's College (see Queen's: The First Hundred and Fifty Years
(Newburgh: Hedgehog Productions Inc., 1990), pp. 28-29).
My 1997 enquiries led to the discovery
that Professor George Romanes and his family occupied part of the block from 1846 to 1850
(see M. Angus, "Queen's College on William Street",
Historic Kingston 34 (1986)
86-98 and photo of a painting below
dated 1914). Since George John Romanes was born in 1848, it seems
likely that he was born, and had passed his first two years, but a
few yards from where these words are being typed!
|
|
| 51 I thank Geoffrey S. Smith
(Department of History, Queen's University), and Charlotte and Ruth Forsdyke for
most helpful editorial comments. The painting of 207
William Street by Jane Redpath Drummond (1916) was exhibited at the Agnes
Etherington Arts Centre, and the above photograph (courtesy of Dorothy Farr) is displayed with the permission of Mr. Douglas Petty (owner of the original
painting). |
Embryo Transfer
On 27th April
1890, Walter Heape (1855-1928) successfully transferred rabbit embryos
from one mother to another, thus performing what we believe was the
first mammalian embryo transfer. He thus achieved what Romanes had
attempted but failed to do; namely, to demonstrate that, at least
during the period of embryogenesis, the postulated "gemmules"
or "pangens" of genetic
information postulated by Darwin, could not be transferred from the
mother to the foetus. The new born rabbits had all the characters of
the biological parent. However, while Heape asked "What
effect, if any, does a uterine foster-mother have upon her
foster-children?", he made no claim to be testing Darwin's
hypothesis of pangenesis.
Biggers (1991) suggests
that "Romanes abandoned his ovarian
transplantation and embryo transfer studies when he became convinced,
somewhat surprisingly, that, when two breeds of rabbit hybridize, no
intermediate between the two parental forms are produced."
In theory, at least some of Darwin's "gemmules"
should have transferred between the two parental genomes to produce
intermediate types. Thus, in his own way, Romanes had rediscovered
some of what Naudin and Mendel had discovered in the 1860s.
Romanes did not easily give
up on the possibility of "gemmule"
transfer, which might explain the acquisition of acquired properties (Lamarkism).
Shortly before his premature death in 1894 he wrote to Sharpey-Schafer
(May 18th) who had reported some success in transfer experiments:
"I have found, after
several years experimenting with rats, rabbits and [ ],
that one may breed scores and hundreds of first crosses
between different varieties, and never get a single mongrel
throwing off intermediate characters - or indeed any
resemblance to one side of the house. Yet, if the younger are
subsequently crossed inter se (i.e. brothers and sisters, of
first crossings) the crossed parentage at once repeats itself.
Ergo, even if the pups wh. are born appear to give a negative
result, keep them to breed from with one another." |
Unknown to Romanes (although he had
cited Mendel in a review for the Encyclopedia
Britannica), this is what Mendel reported with pea
crosses in 1865.
Much like Romanes, Walter Heape was independently
wealthy and was educated by a private tutor. He was interested in the
application of scientific knowledge to human welfare, but appreciated
how inadequate was the knowledge base. He supervised the construction
of the Plymouth Laboratory under the auspices of the Marine Biological
Association to serve the fishing industry. At Cambridge, he was a
student of Foster, and here he probably first encountered William
Bateson. At a critical point, he was appointed to the Evolution
Committee of the Royal Society of London. Here, the knives were out
between Bateson and the "Biometricians" (including Weldon).
The Committee ended in backing Bateson. Heape's work was confirmed by
Castle & Phillips (1909) with guinea pigs.
A related issue was whether prior fertilization by one male could be
evident in the offspring of fertilization by another male ("telogony").
Thus, a woman might have, by a second husband, children who resembled
a former husband. The "gemmules"
of the ex-husband would have persisted long enough to influence
later children (a handy excuse for a randy ex). This was investigated by another member of the
Evolution Committee, J. Cossar Ewart (1851-1933),
who had collaborated in research with Romanes in Scotland in the
1870s, and had been supported by Romanes in seeking (and obtaining)
the Chair of Natural History at the University of Edinburgh. He
prompted Lord Rosebery to sponsor a series of lectures by Romanes in
1888 on "The Philosophy of Natural History,"
which formed the basis of the latter's three volume series Darwin,
And After Darwin.
Biggers, J. D. (1991) Walter Heape, FRS: a pioneer in reproductive
biology. Centenary of his embryo transfer experiments. J.
Reprod. Fert. 93, 173-186.
Castle, W. E. & Phillips, J. C.
(1909) A successful ovarian transplantation in the guinea-pig and its
bearing on problems in genetics. Science
30, 312-313. Romanes,
G. J. (1894) Sharpey-Shafer correspondence. Welcome History
of Medicine Museum, London. Acknowledgement.
Dr. K. J. Betteridge provided helpful information on Heape and Ewart.
|
Struggle to Define and Show Relationships between the Four Black Boxes:
1. Variation
2. Heredity
3. Phenotypic ("Natural") Selection/Isolation,
- 4. Reproductive ("Physiological")
Selection/Isolation
Hybrid Sterility (Darwin 1862)
 Variation (Hooker 1862)
Variation (Hooker 1862)
Pangenesis (Darwin 1868)
Inutility of Characters (Gulick
1872)
Natural and Artificial Selection
(Belt 1874)
Inutility
of Characters. Paradox of Sex. Random Drift (Delboeuf 1877)
Struggle to
Establish "Physiological" Selection/Isolation as the most
Important Form of Reproductive Selection/Isolation

An Unnoticed Factor in Evolution
(Catchpool 1884)
Romanes (1886)
Romanes (1887)
Embryo Transfer (Heape
1890)
Struggle to Establish the
"Chromosomal" (not "Genic")
basis of Physiological Selection/Isolation
Hybridism and the Germ-Cell (Guyer
1900, 1902)

Cytological Basis for the
Mendelian Laws (Cannon 1902)
Chromosomes of the Germ Cells
(Montgomery 1901)
Chromosomes in Heredity (Sutton 1903)
Two Levels of Genetic
Information (Bateson & Saunders 1902)
Heredity and Variation in Modern Lights
(Bateson 1909)
A Phenomenon of Arrangement
(Bateson 1914)
Chromosomes, Polyploidy and Why Sex Evolved
(Winge 1917)
The Blind Watchmaker. Review (1987)
Speciation in Retroviruses (1995)
Origin of Species (1996)
Thinking about Stem-Loops (1998)
Two Levels of Information in
DNA (1999)
Haldane's Rule (2000)
Non-Genic (Chromosomal) Speciation (2003)
Allen, Romanes and Gould (2004)
Chromosomal Speciation: A Reply (2004)
Heredity as Information Transfer (2006)
Positive Selection of Synonymous Mutations Initiates Species Divergence (2007)
Molecular Sex (2007)
Darwin's 'Weak Point' (2010)
Some Evolutionary Terms Used in these Pages
Evolutionary
event. Evolution
results from the accumulation of many evolutionary "events", each involving some
or all of four processes: variation, heredity (inheritance), phenotypic
selection, reproductive isolation. An evolutionary event begins with variation
and is followed by the process of heredity continuing through the generations.
Differential selection
may follow variation.
Variation
(like does not produce like). There is variation among members of a biological species
and the resulting variant members are candidates for
differential selection. Variation
is a process which produces the variant features of variant organisms. These variant
features are themselves often referred to as variations. Thus the word "variation"
must be understood in context either as the process, or as the result of that process.
Some variant features are latent or cryptic (i.e. not immediately observable).
Heredity (like produces like). There is inheritance of a
newly acquired variation by at least some of the offspring of variant members, and
inheritance of that variation, in turn, by some of their offspring, and some of the
subsequent offspring. If a variation is not inherited (overtly or cryptically) then it may
be the result of some environmental influence and is not strictly related to the
evolutionary event under consideration. Thus, increased muscular strength
acquired through exercise would not be transferred to offspring.
Blending
inheritance
(like and non-like merge). Recombination being a rare and random
event, each gene tends to be inherited intact and a corresponding
character may emerge unblended with other characters. Thus, the major factor
determining tallness in peas is stem-length, which is controlled by one
gene. The tall character is inherited without blending. The individual pea
plant is either tall or short. There are no intermediates. However, often
dominance is incomplete, so a blended phenotype appears among offspring.
Thus, all the offspring of a cross between red and white flowering plants
may all be pink in the first generation. Furthermore, numerous characters are multigenic
(non-allelic). Thus, tallness in man
depends on the lengths of many individual bones. It was recognized by
Mendel, Bateson and, later, Fisher, that multigenic characters will blend
in offspring (i.e. a range of intermediates will be observed). Thus, the
extent of phenotypic variation offered to natural selection
will be decreased relative to non-blending inheritance. Blending inheritance will tend to slow the rate of
evolutionary change.
Selection.
The words "selection" and "isolation" have led to confusion. The words have the same meaning, although one may be preferred
in a particular context. Selection is something which can be done when there is more than
one of something. If there is only one of something, then it can be
considered as already
selected (or isolated). In much of the literature, "selection"
often means phenotypic selection; "isolation"
often means reproductive selection.
Phenotypic
selection There is positive selection of
members of the species with advantageous variations, and negative selection of members of
the species with disadvantageous variations. This isolation on the basis of phenotypic
adaptations (positive or negative) is the type of selection with which Darwin was
primarily concerned. In this respect he and others spoke of "survival of the
fittest" and the " struggle for existence". He used the term "natural
selection" to distinguish this form of phenotypic
selection from the artificial selection carried out by the
horticulturalist or breeder. However, reproductive
isolation/selection can also be natural or artificial.
Reproductive
isolation. Variation, heredity and
phenotypic selection can operate so that advantageous phenotypic characteristics
tend to increase in a population, and disadvantageous phenotypic characteristics
tend to decrease in a population. They provide for the adaptation of species, and operate primarily at the level of the individual member
of the species (individual selection). Ultimately, this linear process
(isolation of the fit from the unfit) can
lead to new species (i.e. one that would have been reproductively
incompatible with the ancestral species if the cross were still possible). Here the fit are
reproductively isolated
from the unfit, because the latter are eliminated. However, sometimes an
origin of species arises from a divergence
between two equally fit populations, and reproductive isolation
is needed more directly. To originate species in this way (isolation of the fit from the
fit) groups have to be selected. Speciation
(species selection) is a form of group
selection .
A group is a set of units. For our purposes, if there is only one unit in a set it is an
individual, and does not constitute a group. A group whose members reproduce only
asexually is, by definition, already reproductively isolated from other groups.
Species
. The most secure definition of a biological
species is a group of organisms which is reproductively
isolated
from other groups of organisms. Members of the group are unable to cross
(continue the line) with all
other organisms except those organisms which belong to the group.
Inability to cross includes the production of offspring that are sterile
(infertile). Degrees of infertility
are characteristic of organisms at the species interface (i.e. potential incipient
species).
Hybrid. The offspring of crosses between members of two anatomically distinct
lines (varieties, races) are often referred to as "hybrids". Since the
definition of species is based primarily on reproductive performance, rather than
anatomical difference, this can lead to confusion. For our purposes any offspring is the
hybrid of its parents if the parents are different genomically. There may
be hybrid vigour
("positive heterosis"). There may be "negative
heterosis" manifest as hybrid
inviability (developmental barrier) or hybrid sterility
(gonadal barrier). For humans, the only possible offspring not a "hybrid" would
be the extremely improbable case of a female who inherited absolutely identical sequences
from both parents. In the case of a human male, even if the autosomes were absolutely
identical, the sex chromosomes would differ; so, under our definition, all males are
hybrids.
More usually, the term is used in the context of a specific character
(e.g. height). An organism may be pure (homozygous) for that character
(e.g. corresponding paternal and maternal chromosomes both contain a gene
conferring tallness), or it may be a hybrid for that character (e.g. the paternal
chromosome may contain a gene conferring tallness and the maternal
chromosome may contain a gene conferring smallness). If tallness is
dominant, then the hybrid (heterozygote) may be as tall as organisms which
are homozygously tall.
Genome.
This term might be interpreted by the uninitiated as "gene
home". We talk of "the human genome" or "the genome of the bacterium Escherichia
coli". We usually mean the most obvious form of genetic material, which in these
organisms is DNA. It is true that most of the E. coli genome consists of genes,
but in "higher" organisms there is much DNA which is not so obviously
"genic". Our genomes are not just "gene homes". Note also that a piece
of the genome which we recognize as a "gene" may be serving functions other than
those conventionally considered "genic". This is considered more in the
Bioinformatics/Genomics section of these pages.
Further Acknowledgements. The
photographs of Romanes and Gulick are from Romanes' Darwin, and After Darwin
(Longmans). Photographs of the
portraits of George and Ethel Romanes were made available by Giles J. Romanes. The
photographs of Charles Darwin and Thomas Huxley are from the History of Medicine Division
of the National Library of Medicine, USA (Click here).
The photographs and drawing of Bateson are from the Report of the Royal Horticultural
Conference on Genetics, 1906, as reproduced by H. F. Roberts in Plant
Hybridization Before Mendel (Princeton University Press, 1929), and Beatrice
Bateson's two books Letters from the Steppe (Methuen, London, 1928), and William
Bateson F.R.S. (Cambridge University Press, 1928; original in the National Portrait
Gallery, London). In the Delboeuf paper, the beautiful photograph of a South American
butterfly (Heliconidae) is from Dr. William T. Hark's Butterfly Web Page (Click here), and the orangatan is
from the Web Page of the Orangatan Foundation International, Los Angeles. Other
acknowledgements are given in individual pages. The pictures of gametes and early
divisions of the zygote in the legend to Figure 1 are from the prolife webpage of
Simbahayan Sa Maynila, who, in turn, acknowledges Dr. Anna E. Ross of Christian Brothers'
University.he
portraits of George and Ethel Romanes were made available by Giles J. Romanes. The
photographs of Charles Darwin and Thomas Huxley are from the History of Medicine Division
of the National Library of Medicine, USA (Click here).
The photographs and drawing of Bateson are from the Report of the Royal Horticultural
Conference on Genetics, 1906, as reproduced by H. F. Roberts in Plant
Hybridization Before Mendel (Princeton University Press, 1929), and Beatrice
Bateson's two books Letters from the Steppe (Methuen, London, 1928), and William
Bateson F.R.S. (Cambridge University Press, 1928; original in the National Portrait
Gallery, London). In the Delboeuf paper, the beautiful photograph of a South American
butterfly (Heliconidae) is from Dr. William T. Hark's Butterfly Web Page (Click here), and the orangatan is
from the Web Page of the Orangatan Foundation International, Los Angeles. Other
acknowledgements are given in individual pages. The pictures of gametes and early
divisions of the zygote in the legend to Figure 1 are from the prolife webpage of
Simbahayan Sa Maynila, who, in turn, acknowledges Dr. Anna E. Ross of Christian Brothers'
University. |
Other Web Sites
on Evolution with a Historical Perspective
Darwin
correspondence:
calender, copy of each letter, and much more
Darwin's other
works online:
A complete collection of everything he wrote.
Darwiniana by Asa Gray, 1876:
full-text of book on-line
Darwiniana Essays
Encyclopedia of Life Sciences [See Biographies of Mendel, Romanes, Bateson, Muller, Haldane and Chargaff]
Electronic Scholarly Fascimile Project on Classical Genetics
Electronic Scholarly
Fascimile of "Mendel's Principles of Heredity. A Defence". By W.
Bateson
Fisher,
Ronald A, - Papers:
University of Adelaide
Galton, Francis ["Biometrician"]
Gould, Stephen Jay [Advocate of hierarchical levels]
Grant Allen
[born Kingston
1848]
Hooker, Joseph Dalton
[Proposer
of "Creation
by Variation"]
Huxley,
Thomas Henry [Darwin's "bulldog"]
Korthof's Reviews of Books on Evolution
Mallet's History of Species Concepts
MendelWeb
Mivart,
St. George on "The Genesis of Species" 1871
Panspermia [An unlikely hypothesis, but a lucid
discussion of the issues.]
Wallace, Alfred Russel
[Search Index of Charles Smith's webpages on Alfred Wallace
for my contributions]
Books on Evolution
Videos on Evolution for Beginners
Return to Home Page |
|