|
|
|
|
|
|
|
|
This
"preprint version" differs from the "definitive published version"
in the Notes
and Records of the Royal Society (2010) 64, 139-154, doi:10.1098/rsnr.2009.0045
Click Here
The latter was posted as "Online FirstCite Alert" on 29th
October 2009 and, to celebrate the 350th annniversary of the RS, with other
papers was made freely available (open access).
|
"History is rich in examples of disregarded ideas --, some of them of an importance that came to be recognized long after their publication. One such instance is discussed in Donald Forsdyke's article on the theory of 'physiological selection' that George Romanes advanced in 1886 and William Bateson subsequently championed as a way of explaining the failure of breeders to produce sterile hybrids from intra-species (as opposed to inter-species) crosses. The Romanes-Bateson hypothesis found little support for more than a hundred years, but now, as Forsdyke observes, it is receiving favourable attention in the light of modern work on yeast hybrids." Robin Fox, Editor |
Based on an address - "Bateson's contributions to evolutionary theory" - delivered at the
John Innes Centenary Symposium (9th September 2009), and at
the Galton Institute Conference on "William Bateson: his
Exceptions and the Origin of Species Revisited" (The Royal Society, 1st
October 2009). Online versions of the addresses are at the websites of
the John
|
![]()
BATESON APPROVED "AN ADDITIONAL SUGGESTION"
CRYPTIC VARIABILITY OF THE REPRODUCTIVE SYSTEM
COLLECTIVE VARIATION OF A POPULATION SUBSET
SPORADIC AND REGULAR STERILITIES
CHROMOSOMES AND THE RESIDUE OR IRRESOLUBLE BASE
TWO FACTORS MUST COMPLEMENT TO PRODUCE STERILITY
COMPLEMENTATION LIKE SWORD AND SCABBARD
End_Note_(Feb_2015)__Origin_of_the_term_residue
![]()
|
SUMMARY Huxley
identified as In his theory of "physiological selection," Romanes postulated germ-line "collective variations" that accumulate in certain members of a species, who are thus "physiological complements" producing fertile offspring when mutually crossed, but sterile offspring when crossed with others. Unlike Darwin's natural selection that secured reproductive isolation of the fit by elimination of the unfit, physiological selection postulated variations in the reproductive system that were not targets of natural selection; these sympatrically isolated the fit from the fit, leaving two species where there was initially one. Bateson approved physiological selection. He noted that Mendel's "unit characters" were "sensible manifestations" of what we now refer to as "genes," but postulated a "residue," distinct from genes, that might affect gene flow between organisms and so originate species. The reproductive isolation of the parents of a sterile hybrid was due to two complementing non-genic factors (the "residue") separately introduced into the hybrid by each parent. Modern studies, especially of yeast hybrids, support the Romanes-Bateson viewpoint. |
![]()
|
Many organisms repeat the cycle adult-gametes-zygote-adult through the generations. When lineages diverge this cycle is interrupted and two separate cycles result. The interruption - a potential origin of species - can occur at any stage of the cycle, either prezygotically when gametes meet to form a zygote, or postzygotically when the zygote develops into an adult that can then form gametes. Since each stage requires the action of specific gene products, it follows that changes in certain genes might lead to cycle interruption and divergence into two reproductively isolated lines [1]. However, "speciation genes" have been hard to identify [2], and a classical example has recently been questioned [3]. The purpose of
this paper is, not to question the genic view, but to remind readers of a non-genic
view suggested in 1886 by George Romanes, who was Charles Darwin's research
associate for eight years prior to the latter's death in 1882. This view was
further developed by William Bateson, who in 1900 brought Mendel's now famous
1865 paper to the attention of the English-speaking world. In particular,
Romanes and Bateson noted what Thomas Huxley had referred to as "the weak
point" in Darwin's theory [4]. If offspring could be obtained from
an inter-species cross they were usually sterile (hybrid sterility), but no
breeder had ever produced new forms from intra-species crosses that could be so
characterized. Furthermore, in proposing that evolution proceeded smoothly
without jumps, punctuations or discontinuities, Whereas the work of Mendel went unnoticed for a mere 35 years, the views of Romanes and Bateson were disparaged or went unnoticed for over a century. This was due partly to the twentieth century's obsession with genes, but also to the fact that they were struggling with novel terminology. With much of this (e.g. "allele", "homozygote") we are now familiar. But terms such as "collective variation" and "residue," now seen as highly relevant to speciation studies [6], have largely disappeared from view. Proceeding chronologically from the late Victorian era to modern times, I here reintroduce the latter terms and, through textual analyses of selective quotations [7], let Romanes and Bateson retell their story [8]. |
BATESON APPROVED "AN ADDITIONAL SUGGESTION"
|
In the spring of 1886 William Bateson set off on an eighteen month scientific expedition to the Russian Steppes with the intention of catching up with the relevant literature on his return. However, in the autumn his sister Anna, who was then working with Francis Darwin, dispatched three consecutive issues of Nature containing a serialized May address to the Linnean Society. Here Romanes had made an "additional suggestion on the origin of species" that he named "Physiological Selection," or the "Evolution of Species by Independent Variation" [9]. This had raised a storm of criticism from the senior Darwinians (Alfred Wallace, Thomas Huxley, Joseph Hooker) and their junior surrogates (Francis Darwin, William Thiselton Dyer, E. Ray Lankester, Raphael Meldola). But the response of the 25 year old Bateson (Kasalinsk, Dec. 4th, 1886) indicated tempered approval:
Another young English biologist, the 27 year old Joseph Cunningham, was also likely to have approved (see below). |
CRYPTIC VARIABILITY OF THE REPRODUCTIVE SYSTEM
|
Darwin viewed hybrid sterility as a puzzling negative trait that should have been countered by natural selection [11]. In his Linnean address Romanes went beyond this, pointing out that, apart from variations between individuals that we can easily detect (e.g. differences in eye color), there are likely to be variations that we cannot easily detect. Some variations in parental gonads might ultimately be detected by their conferral of the sterility phenotype on the otherwise healthy offspring of certain pairs. This would designate the parents as reproductively isolated from each other. The partner-specific reproductive-isolation-phenotype of the parents would be manifest as the sterility phenotype of their offspring. In Romanes' view, the establishment of this form of reproductive isolation - isolation of the fit from the fit - would in many cases have been critical for branching into two species, with or without the subsequent intervention of natural selection that would isolate (by elimination) the unfit from the fit (linear, non-branching, evolution). Romanes' physiological selection was an abstraction for which there was then no cytological or biochemical basis. To give his Victorian audience an example of one form of physiological selection, Romanes pointed to a variation that might lead a plant to flower earlier than most members of its species. Early flowering forms would be reproductively isolated from late flowering forms by a temporal barrier. At an early stage pollen kept from an early flowering form would still be capable of fertilizing flowers of late flowering forms. Over time, incompatibilities between early pollen and late ovules would randomly arise, and this mutual incompatibility (a further barrier) would eventually define the two forms as distinct species. Today we attribute allochronic and gamete-incompatibility barriers, as we attribute allopatric barriers, to differences in genes. But Romanes did not think this form of variation would have operated generally to originate species:
Romanes felt no obligation to further clarify intrinsic causes: "It is enough for the explanation which is furnished by Mr. Darwin's theory of the evolution of adaptive structures by natural selection, that the variations in question take place; and similarly as to the present theory of the evolution of species by physiological selection" [13]. On the other hand he was quite prepared to agree that this form of variation could proceed continuously as Darwin had proposed for the evolution of adaptations:
|
COLLECTIVE VARIATION OF A POPULATION SUBSET
|
Anticipating modern developments in population genetics, Romanes thought the slow increase of his postulated independent variation of the reproductive system might reflect its "collective" nature, in that it would affect a distinct lineage within a population. One of the misconceptions of his opponents was:
Free-intercrossing would have tended to swamp or blend the sporadically appearing variations upon which natural selection might act. But Romanes' collective "independent variation" of the reproductive system - postulated to be a "general variability - - about a mean" (e.g. as human heights form a normal distribution) - was held to militate against this:
Remarkably, two of Romanes' sternest critics provided valuable guidance on terminology. Thiselton Dyer [17] suggested the term "reproductive isolation." Wallace [18] noted that two individuals who had each become reproductively isolated from members of their main species (with whom they might cross to produce healthy but sterile offspring), might produce healthy and fertile offspring when mutually crossed. Hence the parents could be considered as "physiological complements." Another critic, Huxley, failed to recognize that his "weak point" criticism of Darwin had been met [19]. Romanes had distinguished two types of variation: overt sporadic variations that might lead to the evolution by natural selection of better adapted forms (which the breeder might capture and selectively pair), and cryptic collective variations that might lead to the evolution of species by physiological selection (which the breeder would find hard to simulate). |
SPORADIC AND REGULAR STERILITIES
|
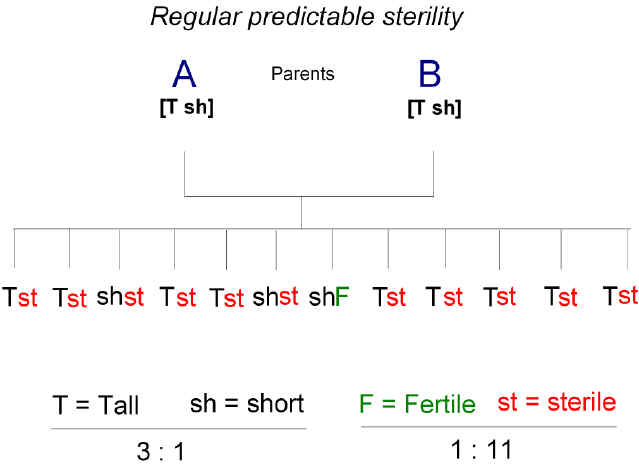
Romanes died at age 46 in 1894, the year that Bateson published a major treatise - "Materials for the Study of Variation, Treated with Especial Regard to Discontinuity in the Origin of Species" [20]. At that time, in keeping with Huxley, Bateson saw the origin of species as a discontinuous process, similar to that manifest as the sudden arising, in one generation, of a new variety of flower with one perfect extra petal. The suddenness of the emergence implied a discontinuous underlying mechanism uninfluenced by natural selection. The book was warmly reviewed by the fish biologist Joseph Cunningham, who noted similarities to Romanes' physiological selection theory, but declared Bateson's argument to be "defective" since there was "no necessity for discontinuity of variation to explain discontinuity of species" [21]. Like various continental botanists (Hugo de Vries, Carl Correns, Erich von Tschermak), in the 1890s Bateson and E. Rebecca Saunders had begun crossing various lines of plants and animals and scoring the inheritance of characters among the progeny. Following the discovery of Mendel's work in 1900, Bateson became its leading advocate in the English-speaking world. Given the intense opposition he faced from the mathematical biologists ("biometricians"), it is not surprising that his attention was directed more to verifying Mendel's laws than to pronouncing on their cellular basis. However, in October 1902 at the Second International Conference on Plant Breeding and Hybridization in New York he noted: "We have reason to believe that the chromosomes of the father plant and mother plant, side by side, represent blocks of parental characters" [22]. In December 1901 Bateson and Saunders had submitted a detailed 160 page report to the Evolution Committee of the Royal Society, to which they made minor amendments in March 1902 [23]. The Report noted that the classical sterility of the hybrids resulting from a cross between species that were believed to have recently diverged from a common ancestor (allied species), did not follow a Mendelian pattern of character assortment among offspring. Although the offspring might have inherited some conventional character, such as color or height, in a Mendelian fashion (e.g. 3:1 ratio), usually the same offspring would all inherit the sterility phenotype (figure 1). At that time they had to admit:
|
|
|
Figure 1. The regular, predictable, sterility observed when members of closely allied species are crossed. Parents, A and B, are heterozygous for height, with an allele for tallness (T) and an allele for shortness (sh). If tallness is dominant, the ratio of tall to short offspring is 3:1, in accord with Mendelian predictions. However, the majority of offspring are sterile (st), not fertile (F). |
|
Thus, the hybrid sterility phenotype that resulted from a cross between allied species was something intrinsic to an organism and did not relate to extrinsic environmental elements. That they were thinking of an imperfection in the pairing of homologous parental chromosomes is indicated by their citing Guyer who "in ignorance of Mendel's work" had independently derived Mendel's laws from the cytology of spermatogenesis in pigeon hybrids [25]. Furthermore, early in 1902 Bateson noted:
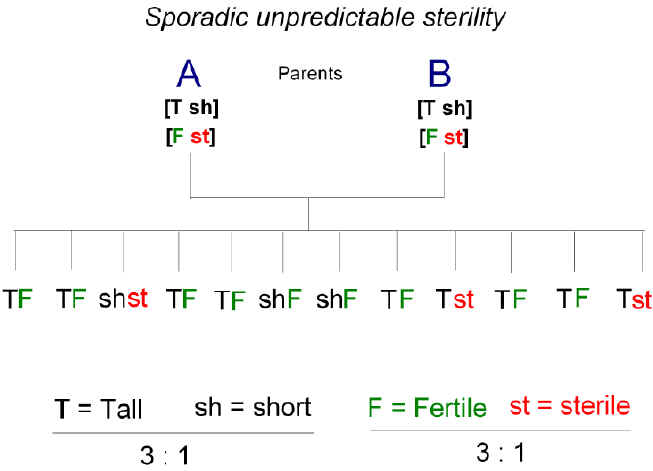
There was repeated reference to Guyer in Bateson's later works [27]. But soon Bateson and his colleagues came across cases of infertility that followed Mendelian patterns of inheritance (figure 2). Their second report to the Evolution Committee in 1905 [28] drew a clear distinction between this unpredictable sporadic sterility seen among the offspring of crosses between closely related individuals, and the predictable sterility seen among the offspring of crosses between distantly related individuals (i.e. members of allied species):
|
|
|
Figure 2. The sporadic, unpredictable, sterility observed among the offspring of a within-species cross. Parents, A and B, are heterozygous for height as in Fig. 1. They are also (unknowingly) heterozygous for a recessive sterility gene (st). The ratios of dominant to recessive among offspring accord with Mendelian predictions, thus indicating that sterility can be of genic origin. |
|
Today we would interpret the latter sterility as a malfunction due to mutation in a gene determining one of the many cell components required for anther development. In this case the component seemed to be required for the reduction division of meiosis. Whatever the ultimate basis of the sterility, Bateson and his colleagues pointed out that this sporadic event arising from a within-species cross (unpredictable in that it had not been anticipated when the cross was first made), contrasted with the regular (predictable) F1 sterility appearing when allied species in general were crossed. Only the former displayed a Mendelian (i.e. genic) pattern of inheritance. Indeed, in one case the sterility character was found to be coupled (linked) to another character displaying Mendelian inheritance (leaf axil color; table 1). |
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
In 1902 Bateson and Saunders recognized that Mendel's unit characters were "the sensible manifestations of physiological units of as yet unknown nature"[30]. Although Bateson did not begin to use the word "gene" until the second decade of the twentieth century, it is employed here when referring to the Mendelian "physiological units" that determine characters. |
CHROMOSOMES AND THE RESIDUE OR IRRESOLUBLE BASE
|
Bateson was among the first to see a link between Mendelian "genes" and chromosomes, and cited Guyer in this context [31]. However, as evidence for a chromosomal location became stronger, Bateson's doubts increased. It was not a question of Bateson's not seeing the wood for the trees. His idea of what constituted the wood differed from that of others. They saw a collection of genes. He saw the genes plus something else that was in some way related to the question of the origin of species. What was the something else and where was it located? Was it on the chromosomes like the Mendelian factors? Or was it elsewhere, perhaps outside the nucleus in the cytoplasm? No clear evidence on this had emerged by 1926, the year of Bateson's death. Thus his pronouncements on chromosomes were always carefully hedged [32]. That the story
was somehow incomplete was made explicit early in 1902: "Has a given organism
a fixed number of unit characters? Can we rightly conceive of the whole organism
as composed of such unit characters, or is there some residue - a basis -
upon which the unit characters are imposed?"[33] This was expressed
in slightly different form a few months later: "From the fact of the existence
of interchangeable [allelic] characters we must, for purposes of treatment, and
to complete the possibilities, necessarily form the conception of an irresoluble
base [i.e. a base not resolvable by Mendelian analysis], though whether such
a conception has any objective reality we have no means as yet of
determining."[34] The report of Bateson and Saunders linked the
concept to
|
TWO FACTORS MUST COMPLEMENT TO PRODUCE STERILITY
|
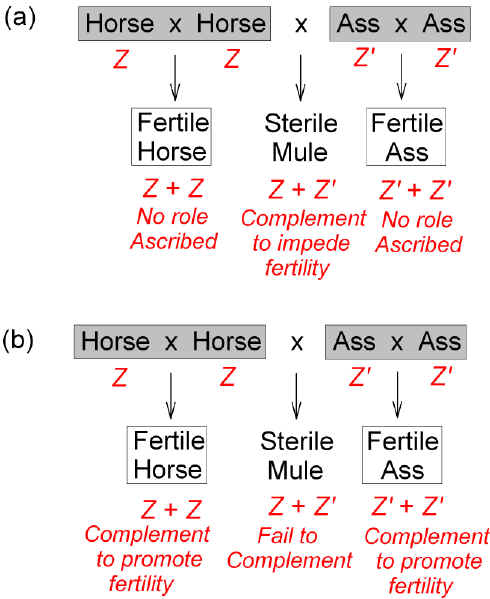
Bateson was considering the sterile offspring of crosses between allied species that were presumed to have derived from a common ancestral species. Since the offspring of crosses between members of that ancestral species would have been fertile (otherwise there would have been no descendents), then the hybrid sterility revealed the emergence of a reproductive barrier between the descendent species (i.e. an interruption of the unitary generational cycle). This could have been an originating barrier leading to the state of reproductive isolation that defined them as distinct species. With little indication that he was writing about chromosomes, the residue, or an irresoluble base, Bateson developed this theme abstractly in an Essay he contributed to the Darwin centenary celebration in 1909 [36]. Here he concluded that "Failure to divide [to produce gametes] is - - the immediate 'cause' of the sterility," so that "we are justified in supposing that there are factors which can arrest or prevent cell division"[37]. Thus, "When two species, both perfectly fertile severally, produce on crossing a sterile progeny, there is a presumption that the sterility is due to the development in the hybrid of some substance which can only be formed by the meeting of two complementary factors - -"(figure 3a). Applying this line of reasoning to what would appear to be a single species, Bateson continued:
Thus, the factors (say Z') which distinguished certain members of a species from their fellows (bearing say Z) would be latent. They would become patent, revealing a sterility-of-offspring phenotype, only if, by chance, there were a cross between certain individuals that had come to differ in the factors (Z' and Z; not Z' and Z', or Z and Z). At least one of the individuals would be rare, since the normal abundant types were generally fertile when intercrossed (i.e. no role was ascribed to Z in the presence of Z, or Z' in the presence of Z'). How often could this happen?
Sterility-of-offspring being a parental phenotype, the fact that a completely sterile offspring could not be subjected to further breeding analysis would not matter. The parents of the offspring complemented each other sufficiently to produce the phenotype, but the complementary factor of one parent (say Z') might be more prone to produce the phenotype than the complementary factor of the other (say Z). Through segregation (further cross-breeding of each parent with other individuals) these complementary factors might be investigated. |
|
|
Figure 3. Hybrid sterility in crosses between allied species (e.g. horse, ass) as due to hypothetical factors, Z and Z', which either (a) complement to bring about a negative effect, or (b) fail to complement to bring about a positive effect. In both cases the result is negative (i.e. sterility). Since parents display fertility with their own kind (intraspecies cross), in 1909 Bateson deduced (a) that in an interspecies cross Z and Z' complement to bring about a negative effect. In 1913, Bateson entertained the converse (b), that Z and Z' fail to complement to bring about a positive effect. |
|
In his 1909 Essay, Bateson considered two sterility factors working together (complementing) to impede fertility (figure 3a). However, he later noted that organisms could be reproductively incompatible because the reproductive system of "each is lacking in one of two complementary elements" that promote fertility (figure 3b) [40]. This view, reminiscent of Romanes, was later adopted by Richard Goldschmidt, who proposed that normally parents contribute complementary factors (e.g. Z and Z) making parental chromosomes compatible at meiosis in their hybrids, which hence are fertile (i.e. the parental factors work together to produce a positive effect). When the factors are not sufficiently complementary (e.g. Z and Z') the parental chromosomes are incompatible in their hybrids, which hence are sterile [41]. |
COMPLEMENTATION LIKE SWORD AND SCABBARD
|
Bateson persisted with his Huxleyan idea that the sterility of a hybrid
resulting from a cross between allied species was "a distinction in kind, of a
nature other than those we perceive among our varieties" [42]. And he was
still attached to the Huxleyan idea of discontinuous variation. In December 1921
he gave his famous address - "Evolutionary Faith and Modern
Doubts" - at the
|
|
|
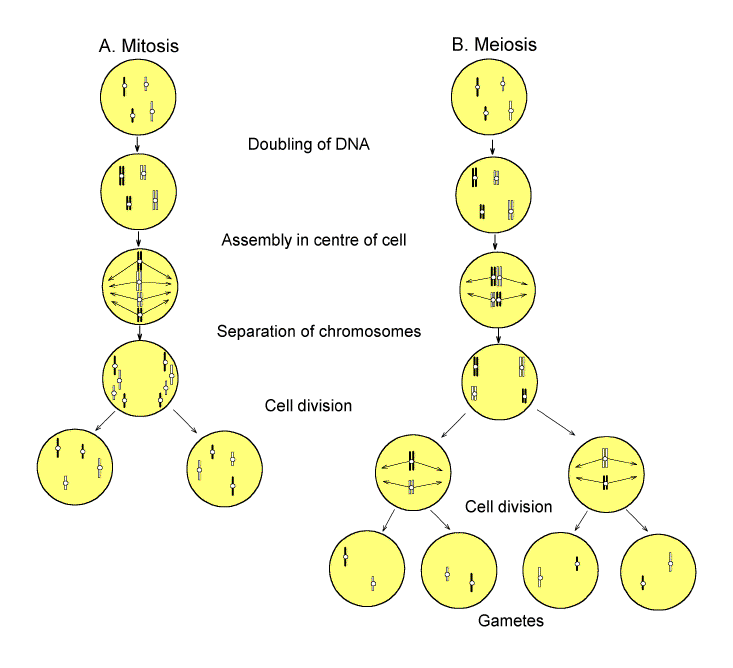
Figure 4. Outline of mitotic divisions that occur in somatic cells (a) and meiotic divisions that occur in cells of the germ-line (b). The chromosomes of the maternal and paternal parents are white and black, respectively. Pairing ("conjugation") of homologous chromosomes occurs in meiosis. |
|
A subsequent interchange with C. R. Crowther is of particular interest [44]. Questioning Bateson's attitude to the mechanism of meiotic chromosome pairing (figure 4), Crowther began by noting that, while parental chromosomes had to cooperate for development of the zygote from embryo to adult, a far higher degree of cooperation would be needed when the chromosomes paired (conjugated) in the gonad of that adult:
Bateson's disparagement of the idea that species might be "a product of a summation of variations" left Crowther "frankly puzzled," for "the proposition is certainly not self-evident." Surely, if the sterility of an offspring were due to a failure within that offspring of homologous chromosomes to conjugate, it mattered little whether the lack of complementarity responsible for that failure was produced by one large variation, or by the summation of many smaller variations. That Crowther was thinking of primary variations occurring at the chromosomal level, rather than anatomical variations of the sterile individual, was explicit:
Bateson conceded that discontinuity of variation was not critical:
|
|
In his 1909 Essay, Bateson did not spell out the relation between his postulated complementary sterility factors and the above "residue" or "irresoluble base." In Bateson's mind the "residue" was not only distinct from genes - the agencies determining the "transferable characters" that were the subject of Mendelian analysis - but it could also limit the extent to which organisms could exchange genes. This limitation (i.e. absence of gene flow) would be complete when the organisms belonged to independent species:
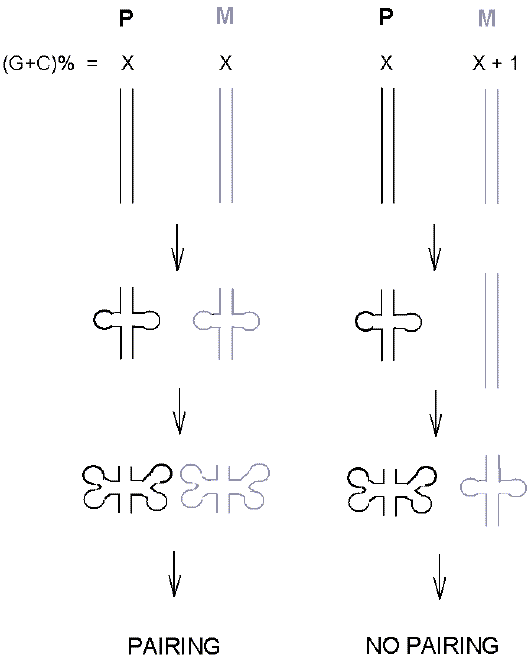
Bateson agreed with Crowther that a fundamental form of reproductive isolation, manifest as the hybrid sterility seen when allied species were crossed, was due to an incompatibility that could be characterized cytologically as impaired pairing of paternal and maternal chromosomes at meiosis. It was inferred that if we can understand what makes chromosomes incompatible, then we can understand hybrid sterility. And if we can understand hybrid sterility, we can understand an origin of species. But how do chromosomes that are homologous (i.e. are alike) pair with each other? Do they pair by virtue of this likeness (like-with-like), of by virtue of some key-in-lock (sword-in-scabbard) complementarity, which implies that they are not really alike? One must be the sword and the other the scabbard. This paradox was resolved when it was appreciated that hereditary information was stored and transmitted as duplex DNA with two strands - a "Watson" strand and a "Crick" strand - that paired with each other by virtue of base complementarity [47]. So, in Crowther's terminology, potentially the sword strand of one chromosome can pair with the scabbard strand of the homologous chromosome (and vice versa). For this purpose, swords have to be unsheathed from their own scabbards and then each inserted into the scabbards of the other. Thus the Watson strand of one chromosome can pair with the Crick strand of the other, and vice versa. This requires that the Watson strand be displaced from pairing with the Crick strand of its own chromosome. Likewise, the Crick strand of the homologous chromosome must be displaced from pairing with the Watson strand of its own chromosome. Then cross-pairing (homology search without strand-breakage) can occur, as Crick himself proposed [48]. The pairing requires complementarity of DNA base sequences. A sporadically appearing change in a certain base could, if dominant, introduce a new phenotype (e.g. an extra petal), but would not greatly affect the overall complementarity between parental chromosomes. But, over time, base changes, including some affecting the classical phenotype, could accumulate. Romanes' "collective variation" that would build up "in a whole race or strain" can now be interpreted as the variation of the base composition of DNA about some characteristic value for the species (i.e. a normal distribution). When differences between chromosomal homologues reached a critical value, meiotic pairing would be impaired. A possible mechanism for this is described elsewhere [49], and summarized in figure 5. Indeed, assuming experiments with laboratory strains to be relevant to natural yeast populations, such "simple sequence divergence" appears to be the predominant form of species initiation in yeast [50]. The initiation might later be reinforced by segmental chromosomal changes that, in some circumstances, could themselves be capable of initiation, as described elsewhere [51]. |
|
|
Figure 5. Model for the mechanism by which homologous maternal and paternal chromosomes pair on the basis of sequence similarity during meiosis. The two complementary strands of paternal (P) and maternal (M) DNA duplexes, represented as paired vertical lines (top), have inward-looking nucleotide bases (not shown). So a strand in a black P duplex is unable to recognize a complementary strand in a grey M duplex (and vice versa). According to Crick's unpairing hypothesis, when the strands unpair (symmetrically extruded loops), the cross-recognition (pairing) can occur. However, the tempo of unpairing is critically dependent on local base composition (G + C expressed as a percentage of the total bases in the region). When composition values are closely identical (left), then pairing can occur (initiated by loop-loop interactions). A small difference (right) desynchronizes the unpairing so no cross-pairing can occur, meiosis fails, and no gametes are produced. |
|
In the twentieth century the writings of Romanes and Bateson on the origin of species were either disparaged or, more usually, ignored [52]. However, we can now see them as latter-day "Mendels" whose recognition of the non-genic nature of the hybrid sterility arising from crosses between allied species anticipated by a century modern developments in genome analysis. While genic differences differentiate species, the spark that originates species can be non-genic. Bateson's non-genic "residue" is not in the cytoplasm, but is "attached to or implanted upon" the chromosomes where the genes reside. Here mutations corresponding to the "collective variations" of Romanes can accumulate, eventually to exceed a critical value in a population subset (the difference between Z and Z' in the terminology of figure 3). In this event a major discontinuity - an origin of species - can result. We can now see how this might have come about at the DNA level. Further questions - whether, in the general case, genic changes have sparked species originations more often than non-genic changes, and whether the latter have involved multiple base changes more frequently than gross karyotypic changes - are not argued here. The point is that, until controverted, the Romanes-Bateson hypothesis deserves an at least an equal place at the table with other hypotheses. |
ACKNOWLEDGEMENTS
The invitations of the Organizing Committee of the John Innes Centenary Symposium (K. Roberts, C. Lamb, D. Hopwood, E. Coen, S. Wilmot, P. Nurse, W. Bodmer), and of the organizers of the Galton Institute Conference on William Bateson (T. M. Cox and M. Keynes), are greatly appreciated. The recent decease of Milo Keynes is sadly noted. Queen's University hosts my web-pages where full-text versions of some of the cited papers, including the relatively inaccessible texts of Guyer, may be found.
|
NOTES
|
![]()
I thank Adam Wilkins, the Editor of the "Perspectives" section of Genetics, who drew an important reference to my attention (note 3), and made several suggestions for improvement (e.g. putting "Darwin's weak point" in the title). But on the advice of two reviewers (the first of whom also made valuable suggestions), he declined (July 2009) to accept the revised paper (first submitted Feb 2009) for publication in Genetics. Since there was an invitation to present the work at the Royal Society in October, the paper was then submitted to Notes & Records of the Royal Society (6th July). There were two prompt positive reviews, one of which observed:
The comments of Wilkin's second reviewer (for Genetics) indicate the gulf that remains to be bridged:
While the present paper was being declined, three papers were being accepted for inclusion in a November commemorative issue of Genetics "Celebrating The Origin of Species". These were by Brian and Deborah Charlesworth ("Darwin and Genetics"), by H. Allen Orr ("Darwin and Darwinism"), and by Adam Wilkins ("Charles Darwin: Genius or Plodder"). [Subsequently, Orr joined Wilkins as coeditor of the "Perspectives" section of Genetics. DRF Aug 2011.] While noting Darwin's puzzlement over the apparently non-adaptive nature of hybrid sterility, the Charlesworths noted:
It is of note that Romanes presented his interpretation of reproductive isolation in 1886, albeit only in general terms. The above model (Figure 5) seems at odds with contemporary models that propose strand breakage and then homology search with single strands. However, the latter models do not exclude the possibility of a search involving initial "kissing" interactions between loops formed by such single strands. |
The doubts cast on genic speciation hypotheses by Louis and Greig, based on their yeast studies (note 2), are supported by work from the Sherlock laboratory at Stanford University:
Kao KC, Schwartz K, Sherlock G (2010) A genome-wide analysis reveals no nuclear Dobzhansky-Muller pairs of determinants of speciation between S. cerevisiae and S. paradoxus, but suggests more complex incompatibilities. PLOS Genetics 6, e1001038. |
According to Vorzimmer (1970), Wallace in his early writings (letter to Darwin February 1868) found a way round Darwin's "weak point" by looking at the issue from the perspective of the line of organisms ("form") rather than of the sterile individual itself:
Vorzimmer notes: "One can see that the fundamental difference between Darwin and Wallace lay in the fact that the former based his utilitarian principle of selection on the basis of the individual organism, while the latter thought in terms of the species" [i.e. "form"]. Vorzimmer PJ (1970) Charles Darwin. The Years of Controversy. Temple University Press, Philadelphia, pp. 206-207. |
An equal place at the table of evolutionary discourse has been accorded by Eviatar Nevo (2012):
This is species selection - one form of group selection - which Wallace acknowledged when writing of selection acting on "the form." Here, due to 'molecular changes,' the selection acts internally after particular pairs of parents have selected each other (externally) to produce offspring (within which the internal selection will act, by virtue of fundamental differences between those particular parents - i.e. they belonged to different groups within what appeared to be a unified species). Nevo E (2012) Speciation: Chromosomal mechanisms. Encyclopedia of Life Sciences. John Wiley, Chichester. |
Following up on the above (Vorzimmer 1970), Darwin turned the discussion over to his mathematical son, George, whose attempts to get through to Wallace tended to cloud rather than clarify. Noting Charles' "failure to be moved" by Wallace's arguments, Vorzimmer thought that, for Wallace, "effective speciation required, not some fortuitous instance of physical [e.g. geographical] isolation - - , but a form of intrinsic isolation such as that resulting through the selective aquisition of sterility." Indeed on 24th March 1868 Wallace wrote to Darwin:
This approximates to the case Romanes, with his "collective variations," was to make in 1886 after the death of Darwin (as set out in these web-pages). Sadly Wallace and Romanes could not see that they were in agreement, and there was some sour correspondence between them circa 1890. Vorzimmer PJ (1970) Charles Darwin. The Years of Controversy. Temple University Press, Philadelphia, pp. 208-209. |
End Note (Feb 2015) Origin of the term "residue"
The term "residue" in the context employed by Bateson appears strange to modern readers. Did he just pull it out of the air, or was it used with a precision that readers at that time would have understood? Here are the opening lines of an essay entitled "The Hidden Self" by the American philosopher/psychologist/physician William James (1842-1910) that was published in Scribner's Magazine (March 1890; 7(3) 361-374), and reproduced in a volume of collected essays in 1897:
Bateson was highly sceptical of psychical phenomena (e.g. telepathy and spiritualists who claimed to be able to make contact with deceased loved ones). But, given the state of knowledge at that time, it appeared to many of his no less sceptical contemporaries that those who wished to study the subject with scientific objectivity should not be dismissed out of hand. This was where James' essay was heading. That aside, these opening remarks have a compelling logic to anyone who knows anything about progress in science, and they council researchers to treasure the "exceptions" that recur from time-to-time in any discipline. Indeed, "treasure you exceptions" is one of the maxims Bateson advised his research associates to follow. If you think this is somewhat far-fetched, you might note that Bateson's neighbour in Grantchester was the philosopher/mathematician Alfred North Whitehead (1861-1947), who had a great respect for William James. With Bertrand Russell, Whitehead produced Principia Mathematica (1910, 1914) and moved in Harvard in 1924. Mike Zundel (2013) writes of the close Bateson-Whitehead relationship:
Zundel, M. (2013). Gregory Bateson. In J. Helin, T. Hernes, D. Hjoerth, & R. Holt (Eds.), Oxford Handbook of Process Philosophy and Organization Studies (pp. 1-15). Oxford: Oxford University Press. |
![]()
Go to: Bateson pages (Click Here)
Go to: The B in "BDM" (Click Here)
Go to: Videolecture based on this paper (Click Here)
Return to: Evolution Index (Click Here)
Return to: Bioinformatics Index (Click Here)
Return to: Homepage (Click Here)
![]()
Established September 2009, posted on the internet 17 Feb 2010, and was last edited on 13 Nov 2020 by Donald Forsdyke