|
|
The
Chromosomes Their Numbers and General Importance by jvind Winge [Comments in square brackets and italics by DRF unless otherwise stated]
|
|
|
|
The
Chromosomes Their Numbers and General Importance by jvind Winge [Comments in square brackets and italics by DRF unless otherwise stated]
|
|
Comptes Rendus des Travaux du Laboratoire Carlsberg (1917) 13, 131-275. Based on Winge's doctoral thesis.
![]()
|
EIGHT CHAPTERS Following an Introduction (pp. 131-134), Winge's paper is divided into eight chapters, and concludes with a Summary (pp. 264-266):
|
![]()
What this paper does not do
Winge does not cite "authorities" directly, but notes (p. 131):
"certain authorities on heredity [?William Bateson, ?Wilhelm Johannsen] themselves oppose the tendency to allow the chromosomes a too direct importance in this respect: i.e. as the seat of genetic disposition."
Furthermore Winge fails fully to recognize the role chromosomes can play in sex determination, noting (p. 208) that:
"the number of those who still adhere to the idea as to the existence of sex-determinative chromosomes is steadily decreasing, and from our present standpoint we may maintain that it is not the heterochromosome which is the primus motor in sexual differentiation, though it may, in several cases, more particularly in insects, be associated with one of the other sex, chiefly the female. It is thus rather a question of correlation between the appearance of the chromosome set and physiological or other conditions which determine the sex of the organism (the "Index hypothesis" of Hcker)."
What this paper does do
However, Winge splendidly summarizes and explores the implications of the pairing of homologous chromosomes in meiosis. This will be summarized on this web page, in the form of quotations from the relevant chapters.
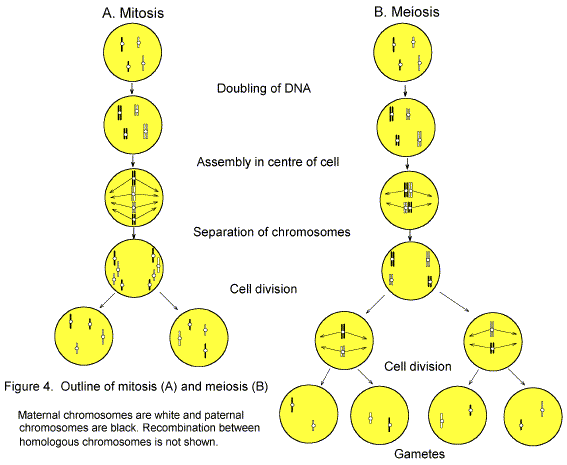
Just to remind you, there is first a modern figure (by DRF) showing (A) normal mitosis (cell division in somatic cells), (B) normal meiosis (cell division, including the reduction division, in gamete-forming cells). The organism is a hybrid between gametes from the father (paternal) and mother (maternal). Since meiosis is normal, the hybrid is fertile. |
|
|
|
We can also note that as
early as 1893, Johann Friedrich Miescher (1844-95),
the discoverer of nucleic acid in gametes and elsewhere, suggested that
sex may have arisen to correct structural defects in molecules:
|
Donald Forsdyke circa 2000
![]()
|
In Chapter 1 Winge notes that " alternation of generations " between " gametophytic " (haploid) and " sporophytic " (diploid) phases is widespread both in animals and plants. Our gametes (one "generation") are haploid and we (the alternate generation) are diploid. The two generations eternally cycle between the two phases. In some forms of life the haploid phase is of longest duration. When Winge refers to an "individual" he often means the organism in this "original form" in the haploid phase of its life cycle (pp. 142-143):
He then extends earlier work of Galton, Butler, Weismann and Montgomery to suggest what we might now refer to as the correction of genetic errors. Winge begins with the simple idea of complementation (pp. 143-145):
|
![]()
Polyploidy and Chromosome Pairing
|
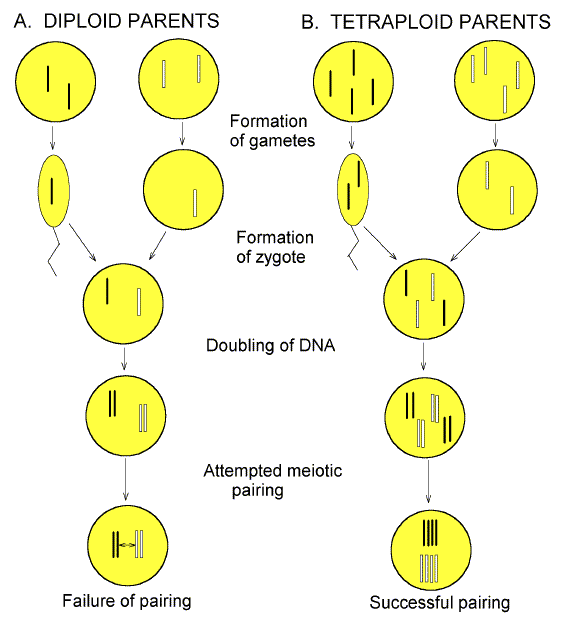
In Chapter 4 Winge examines fertile and unfertile (sterile, apogamous) polyploids, and the pairing of chromosomes, which reveals some "fundamental principle":
The degree of genetic relatedness between two individuals critically affects the pairing of their chromosomes in the hybrid which results from a merging of their gametes:
|
 |
Donald Forsdyke |
|
Winge next offers a classification depending on the "physiogenetic likeness between gametes", a fundamental characteristic determining the number and type of barriers between them:
"1. Philozygoty ["denotes the highest degree of harmony between the sexual cells of two organisms"]... for instance two gametes from the same species of plant. Philozygoty must thus be said normally to characterize gametes belonging to one and the same systematic species [as defined on morphological grounds by biologists]. We may, however, also find that gametes derived from different [again as defined by systematists] species or forms ["heterogamous" as opposed to "homogamous"] have a harmonizing or corresponding "inner physiology" i.e. possess the qualities requisite for a pairing between their chromosomes, and thus the formation of a harmonic common product, a zygote with unimpaired vitality. Heterogamous organisms may thus also be philozygotic. As in the case of homogamous organisms, so also here the parent chromosomes are united in pairs -- at any rate in the gonotokont [gamete forming cell] nuclei, after which they again separate, the two sets of parent chromosomes being distributed on the daughter cells [to become gametes] during reduction division. 2, Pathozygoty. By this I mean ... that two gametes may enter into the formation of a common zygote [i.e. no prezygotic barrier], which, however, owing to the less marked harmony between the constitution of the gametes, may often be only partly capable of development, or not capable of development in the normal manner [hybrid inviability or sterility]. This category will only include gametes derived from different species or races, i.e. heterogamous gametes. When such heterogamous gametes unite, one of the following alternatives must take place:
B. Indirect chromosome union [Polyploidy]. Where a less-marked harmony exists, we must then suppose that this will be visibly expressed by the fact that the chromosomes derived from the two gametes will not unite in pairs at all, but distribute themselves throughout the primary cell of the zygote, as if no dualistic relation of any kind existed. If the chromosomes are to find a partner, then each of the chromosomes in the zygote must divide [thus creating a potential pairing partner], for thus indirectly to produce [the potential for] a union of chromosomes, and we must assume that this is realized in the hybrid zygotes which have any possibility of propagating -- in accordance with what we know from experience as to the behaviour of pairs of chromosomes. The hybrid sporophyte thus produced will then have 4x chromosomes, taking the number from the parent gametes as x. After this, either the chromosome pairs will have the power of further separating by reduction division, transmitting one set of the chromosomes from either parent to each of the gametes -- in which case we have a new hybrid organism with the qualities of a pure species and "double" chromosome number [polyploid]; i.e. containing the sum of the chromosome numbers in the parent species. Or, if the power of reduction has been lost, but the power of continued existence [for that generation] otherwise retained, the result will be an apogamous [sterile] species [individual] with 4x chromosomes.... With regard to the apogamous species, the chromosome number here is ... generally just twice that of the related normally sexed species, and as sexual sterility is itself characteristic of many hybrids, I consider it highly probable that apogamous species are derived from crossings between those normally sexed.... C. No chromosome union. Where the mutual harmony between the two united gametes of heterogamous origin is so slight that their relation becomes almost disharmonious, the interaction between the two organisms will in all probability be of briefer duration and of a less intimate character. This might naturally result in the chromosomes of the two gametes failing to unite either directly or indirectly. I believe I am justified in assuming that the chromosomes of sporophytes capable of sexual development must continually act in pairs, and I must accordingly conclude that when neither direct nor indirect union of chromosomes takes place, then the organism is doomed, and can only exist as an embryo -- possibly with the power of cell division, as long as it is nourished by the mother tissues. 3. Misozygoty [prezygotic barrier]. Gametes of systematically widely differing organisms [defined by systematists using morphological criteria] will as a rule altogether lack the power of uniting in a zygote. Not only is there too little similarity between the constitutions of such gametes; there is moreover no harmony between them, and in consequence, nothing calculated to produce the mutual affinity which is inseparable from all fertilization. The term misozygoty then, I use to designate this lack of mutual harmony; a reciprocal disinclination rendering fertilization impossible." |
![]()
Hybrid Sterility within Incipient Species
|
I n Chapter 5 there is discussion of odd, unpaired, "heterochromosomes", which, in some cases are the sex chromosomes. Under the same heading,Winge considers chromosomes in hybrids between members of allied species, where mispairing may result in free chromosomes. Of significance for the conception of incipient species within the parental species we may note Winge's remark:
|
![]()
Effective Pairing Results in Fertility
|
In Chapter 6 Winge considers the hereditary significance of chromosomes, and the actual chemical basis by which homologs might "repair" each other as a result of meiotic pairing. He notes that:
Winge acknowledges evidence, often involving transmission by the female, that in some cases "cytoplasmic peculiarities" may be responsible for hereditary transmission:
However, the main carriers of hereditary information are the chromosomes:
He then considers in detail the nature of the interaction during chromosome pairing:
Finally, Winge considers the implications of this for understanding the deleterious effects of close inbreeding, and answers the objection, -- if sex is so beneficial, how come the dandelions are doing so well?
|
![]()
Defective Pairing Results in Sterility
|
In Chapter 7 Winge considers chromosome in hybrid organisms which often show partial or complete sterility. The hybrid sterility resulting from "heterogeneous fertilization and all that it involves should be particularly valuable in the study of chromosomes". Like Romanes and Bateson, he draws a distinction between what we now know as "genic" and "chromosomal" causes for hybrid sterility.
Winge then considers chromosomal changes occurring in plant and animal hybrids, such as those described by Federley and Guyer (Click Here; although he only cites Guyer's 1900 abstract in Science).
C. D. Darlington continued this further in "The behaviour of polyploids" (1927 Nature 119, 390). |
Ojvind Winge was born in May 1886 (the same month as Romanes' Linnean Society lecture on physiological selection), and died in 1964. Bateson met him in the 1920s. Waclaw Szybalski worked with him from 1947-49 (see "My Road to Ojvind Winge, the Father of Yeast Genetics" 2001. Genetics 158, 1-6).
Photographs were kindly reproduced by Dr. Morten Kielland-Brandt from The Carlsberg Laboratory 1876-1976. Edited by H. Holter and K.Max. Moller. Pub. Rhodos, Copenhagen, 1976. Copyright permission was given by Dr. Gunver Kyhn for the Carlsberg Foundation. In the book there is a brief biography by M. Westergaard (see also Compt. Rend. Trav. Lab. Carlsberg 34, 1-24; 1965 (Click Here)
![]()
End Note (Aug 2015): Hybrid Sterility Precedes Whole Genome Duplication
Analysis of genome sequences usually provides evidence for one or more ancient whole genome duplications (WGDs). A WGD that generates polyploidy may occur within one organism (autopolyploidy) or result from the fusion of two independent genomes that, when of closely related (allied) species, can be referred to as allopolyploidy. With autopolyploidy, differences between the duplicates will accumulate after the fusion event. With allopolyploidy, differences between the two genomes will already exist and there is often hybrid sterility. To these differences will be added further differences that accumulate after the fusion event.
In an elegant phylogenetic analysis of the genome sequences of various yeast species, Marcet-Houben and Gabaldon (2015; PLOS Biology 13(8):e1002220) were able to distinguish between these possibilities. In agreement with Winge's hypothesis, they found that many differences existed prior to the fusion event. This suggests that WGDs arose in order to "cure" the hybrid sterility that would have resulted. The WGD was of immediate selective advantage since it permitted the line to continue.
![]()
Next: The Origin of Species (1996) (Click Here)
Return to: Guyer Page (Click Here)
Return to: Evolution Index (Click Here)
Return to: HomePage (Click Here)
This page was created circa 2000 and last edited 13 Aug 2015 by Donald Forsdyke