Until recently research upon hybrids has been almost
entirely confined to the more practical problems, such as are connected
with the formation of new or better sorts of plants, and students of
pure science have given little attention to the subject. At the present
moment, however, much interest is manifested by scientists in this
direction, and this awakening interest is largely due to the
republication by Correns and De Vries of Gregor Mendel's experiments and
results. The more recent studies have a two fold nature. They are either
experimental, as the well known work of De Vries, Correns, Webber and
Tschernak, or they are cytological; and each phase of the work has been
undertaken entirely independent from the other.
What are the relations of the cytological to the
experimental researches of hybrids? This question can hardly be answered
at this time, but the possible connection between the two may be pointed
out, however.
Although it is not the purpose of this paper to
set forth the conclusions of the experimenters, for illustration and for
reference, it will be convenient to briefly present those of Mendel.
Mendel studied the behavior of hybrid plants, not
only for the first generation - that of the immediate cross - but in the
following ones as well, and learned
- (1) that in the first generation the hybrid shows the characters
of one parent only, but that
- (2) the characters which were latent in this generation appear in
the later ones, and in such a manner that the plants having the
"dormant" and the "recessive" characters bear a
certain and definite ratio to one another.
In this
connection it need only be said that while the conclusions of Mendel
have been verified by the later researches, they have been found to be
limited in their application to a special type of hybrid, namely, the
Pisum type, while the greater number of hybrids belong to other types.
In
the Pisum type the plants behave as if the bundles of inheritance which
were derived from the parents of the original cross were kept separate,
and were delivered as such to the succeeding generation, and, as if in
fertilization, these were united in all possible proportions. This has
been expressed by the following formulae: A
+ A,
A
+ a,
a
+ A,
a
+ a.
The
result is that a certain percent of the hybrids revert to the characters
of the parents of the original cross. Or, in other words, according to
the laws of Mendel, we might expect that the chromatin derived from the
primitive parents maintained its individuality, and was disposed in such
a manner at the time of the maturation mitosis that the resulting sex
cells were not hybrid,
but pure.
A
portion of this hypothesis has already been demonstrated to be true,
namely, in hybrid ascaris, the chromosomes derived from the
variety bivalens keep separate from the one which is evidently
from the univalens parent. And it will appear presently that an
analogous condition may obtain in other hybrids.
That a hybrid sex cell has chromatin of pure descent - that is, has
chromatin from one parent only, as the second law of Mendel apparently
demands - is yet to be proved, and may be doubted on a priori
grounds, since the wall separating the daughter nuclei is laid down at
right angles and not parallel to the spindle of the dividing nucleus.
However, if it can
be shown that the Pisum type of hybrid, for example, is
associated with the general behavior of the chromatin, as just
suggested, a very important and forward step in explanation of the
nature of this and other types of hybrids shall have been made.
On the other hand, it will be quite as important to demonstrate that
variation in hybrids may take place, even if he sex cells are formed in
a manner identical with those in pure races.
Now
that I have pointed out the possible relation between the experimental
and cytological study of hybrids, I hasten to say that, in reality, up
to the present moment there has been no such connection, since the
cytological work was mainly completed before the experimental was
published; and, further, since the forms which have been turned to
account cytologically have not been studied experimentally. With this I
shall proceed to speak briefly of the cytological work.
This has comprised studies by Guyer on the spermatogenesis of hybrid
pigeons and cannas, by Juel on the spermatogenesis of hybrid Syringa,
and by the writer on spermatogenesis of hybrid cotton. In addition, the
study on Ascaris, already alluded to, and also the structural
study of several hybrid plants by Macfarlane may be included here.
Concerning all the plants upon which cytological studies have been made
it may be said that they are first generation hybrids; that is, they are
the offspring of the original cross.
Guyer reports upon the spermatogenesis of hybrid pigeons, and also
cannas, but since the results are analogous and the conclusions
identical I need only outline the results of his study of the former.
I
understand that Guyer found in the pigeon normal and abnormal
spermatozoa and normal and abnormal maturation mitoses. In the normal
male nuclei the first maturation division showed ring-form chromosomes,
of uniform size, and of half the somatic number. He found, on the other
hand, in the abnormal maturation mitoses that the ring-formed
chromosomes might be of two sorts, large and small, or they might all be
small; and if the latter, they were the same number as those in the
somatic nuclei. In such abnormal nuclei there was a tendency of the
chromosomes to form into two groups, and this localization was taken to
indicate the maintenance by the chromosomes of their individuality. The
general conclusion of the author was, that the
variation noted in the maturation mitoses was likely associated with the
variation of the hybrids themselves.
Guyer had, I believe, like results from his study of hybrid cannas,
except in the canna he found all gradiations between direct and indirect
divisions.
The
writer has studies the maturation divisions in the male nucleus of
hybrid cotton, which was supplied to him by Dr. Webber. The results of
this study may be summarized as follows: Two sorts of maturation
divisions were distinguished, on clearly normal, the other abnormal.
Concerning the former it need only be said that the ring-formed
chromosomes of the first division were of uniform size and of reduced
number. The spindle gave no indication of being double, and was entirely
analogous with that in the pure race. The abnormal divisions were all
direct, but varied in a manner of which is is unnecessary here to speak
further. Transitions between direct and indirect divisions were not
observed.
From
my study I concluded that the normal divisions lead to fertility, and
the abnormal to sterility.
The
remaining purely cytological study is that by Juel on the
spermatogeneses of Syringa rothomagensis. Juel
found abnormal mitoses only.
These were in all stages between direct nuclear divisions and indirect,
save only that the chromosomes did not split in the metaphase.
What is
the significance of Juel's results? It is of interest here to note that the
plant may be wholly sterile, since Focke observes that he has seen no
fruit on it - that is, the abnormalities of spermatogenesis seen by Juel
in the hybrid Syringa lead to sterility.
Thus, Juel's results cause one to ask whether some, if not all, of the
abnormal sex nuclei observed by Guyer in pigeon and canna were not
functionless, and whether the normal divisions alone do not lead to
fertility? Such is apparently the case in the hybrid cotton. If only sex
nuclei that undergo typical maturation mitoses form functional
reproduction cells, it is clear that the variation, in some hybrids at
any rate, must be occasioned by other causes than those of the
irregularities in the splitting, distribution and union of the chromatin
of the sex cells.
Here it may be suggested that an experimental study of such a hybrid as
shows a tendency toward maintaining the individuality of its chromosomes
might be carried on with good results. Such a study should show the
relative sterility of the sex cells, and thus it would supplement the
results derived solely from the cytological investigations. This has not
yet been done, however.
While the cytological data just given do not harmonize completely,
further study may show that they are really not antagonistic, but that
the different hybrids studied should either be placed in different
hybrid classes, or that the "splitting" may not occur in the
first, but later generations.
Thus the leading results which have been obtained by the cytological
study of hybrids may be summarized as follows
- (1) The
establishment of one cause of sterility among hybrids.
- (2) The variation
of the hybrid may or may not be associated with abnormalities of
spermatogeneses; and
- (3) That in certain
cases the chromosomes derived from the original parent tend to
preserve their individuality.
Before closing this brief account of the cytological studies I wish to
speak of the work of Macfarlane on the structure of certain hybrids, a
work published about ten years ago. After giving in detail the minute
structure of several hybrids, this author arrives at his general
conclusion concerning the nature of the hybrid, namely, that it is a
blending of the characters of the parents, each parent contribution
equally to the offspring. The hybrid is thus intermediate in structure
between the parents.
Although it may be at this time premature to suggest a relationship
between the results of studies in the structure and those from
experiment, nevertheless it is interesting to note that practically all
of the hybrids reported by Macfarlane are of the intermediate type of
Correns, and perhaps, therefore, such results as Macfarlane obtained
might thus have been expected. From this possibility is suggested that hybrids
which are not of the intermediate type may have a minute structure which
is likewise not intermediate.
No work on this phase of the subject has yet been done, however.
Finally, as has already been stated in the foregoing summary of the
recent cytological work upon hybrids, this line of research and the
experimental have gone on independently of each other, but it may be
reiterated here that in order to
better understand the causes of the variations in hybrids and that of
the differences in capacity for variability
it is highly desirable that the cytological and experimental work go
hand in hand, that cytological work be done on forms that give marked
experimental results.
|


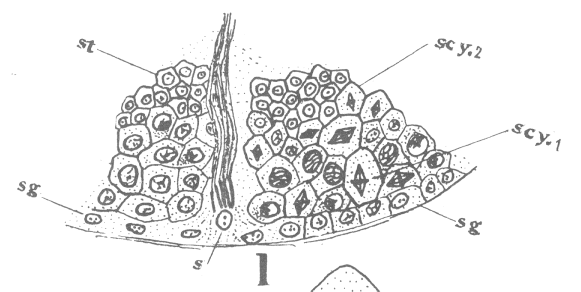
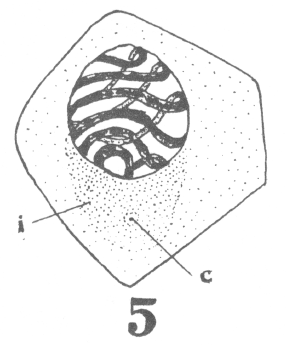
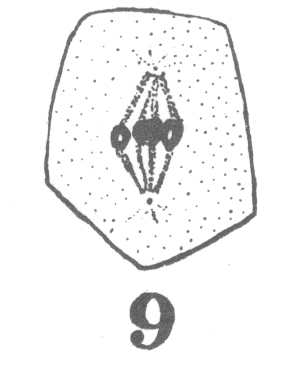
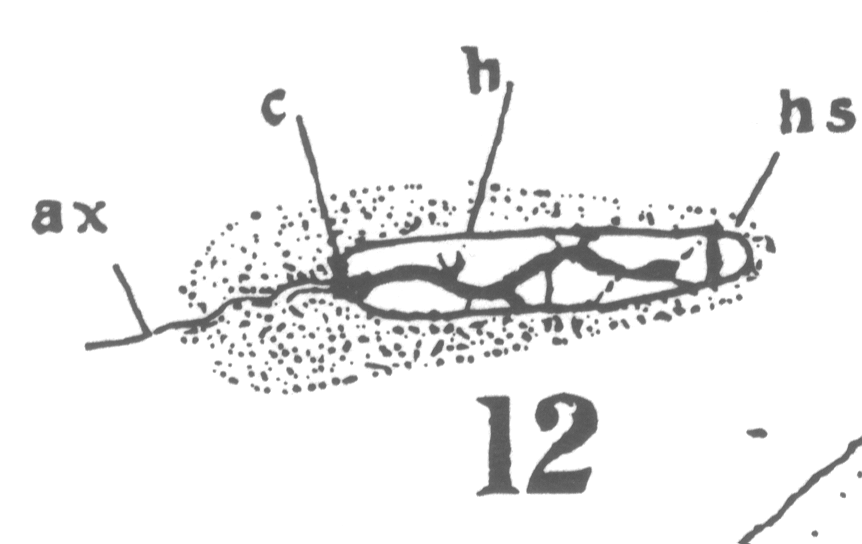
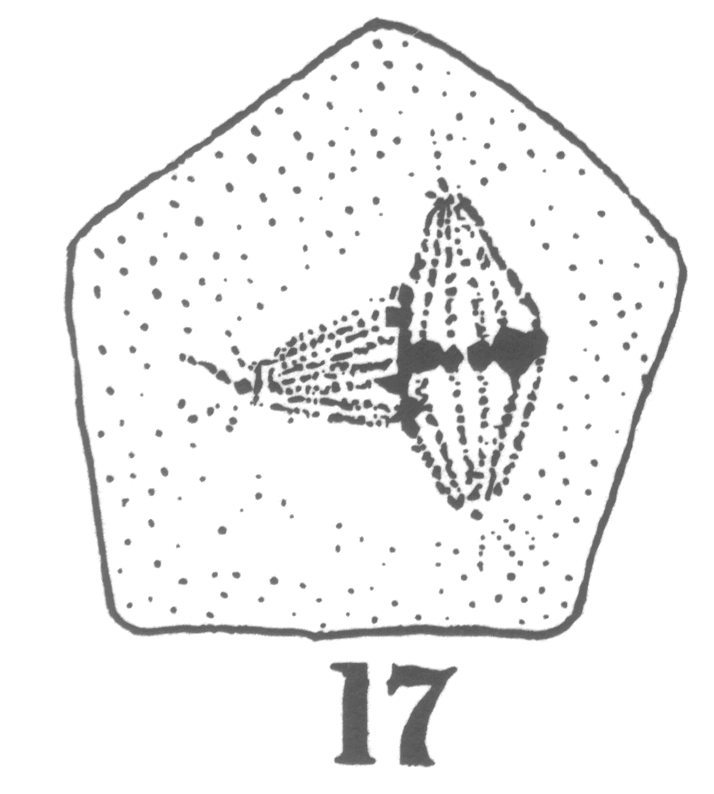
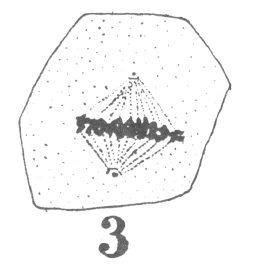 The first
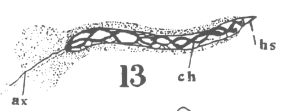
appearance of the spindle fibers is as radiations which spread around the nucleus from the
centrosomes. When the mitotic figure is fully formed, the spindle is short and broad and
the chromosomes lie in a confused band at the equator (Fig. 3; a
spermatogonium ready for division).
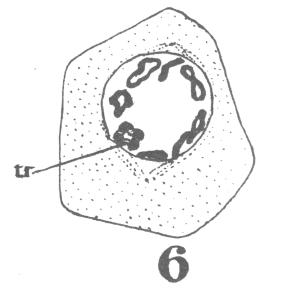
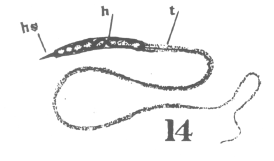
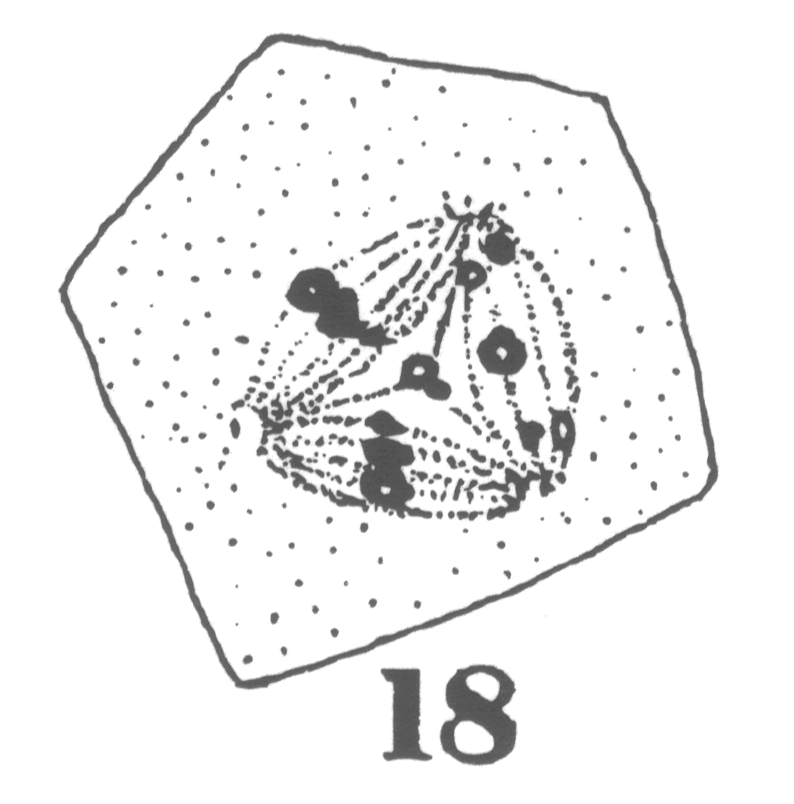
The first
appearance of the spindle fibers is as radiations which spread around the nucleus from the
centrosomes. When the mitotic figure is fully formed, the spindle is short and broad and
the chromosomes lie in a confused band at the equator (Fig. 3; a
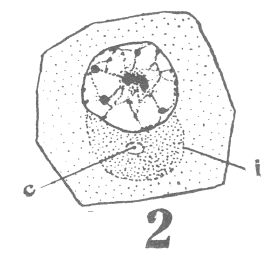
spermatogonium ready for division).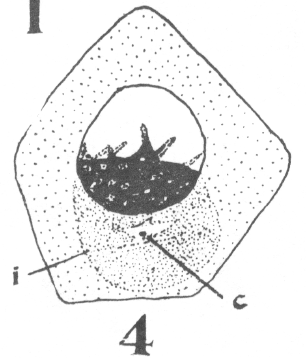 area closely applied to the nucleus, with
the horns of the crescent so extended as to
inclose more than half of the nuclear surface.
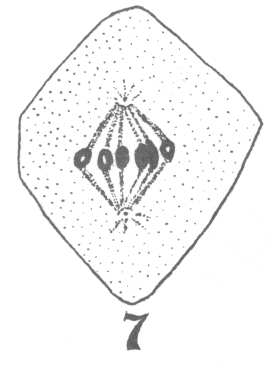
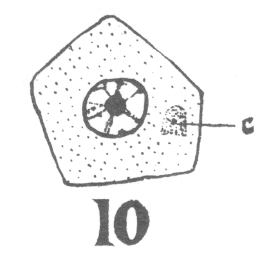
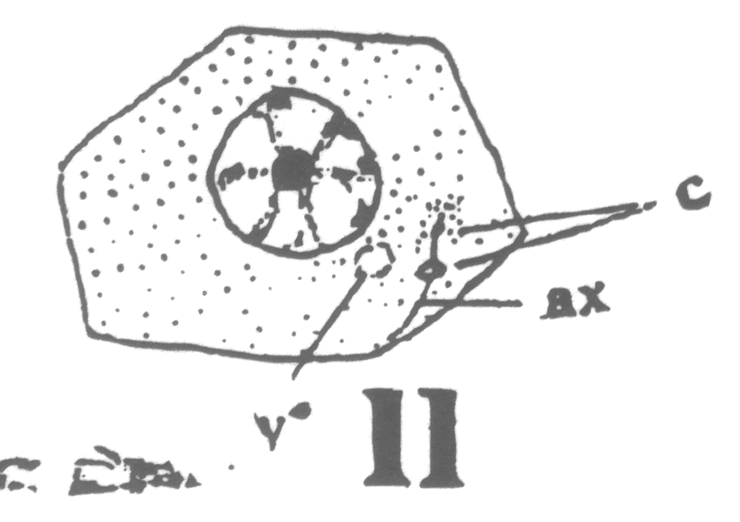
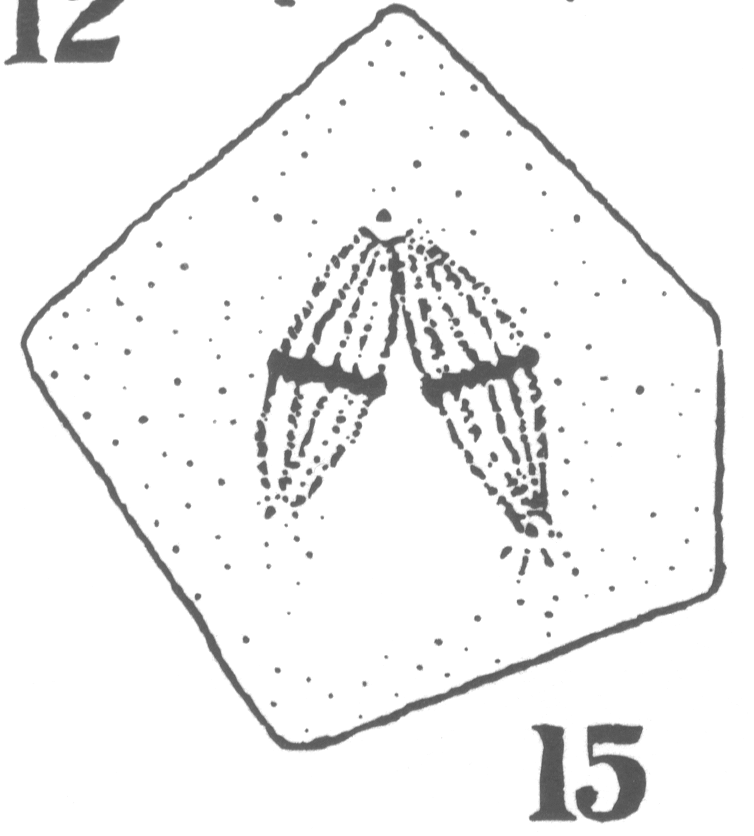
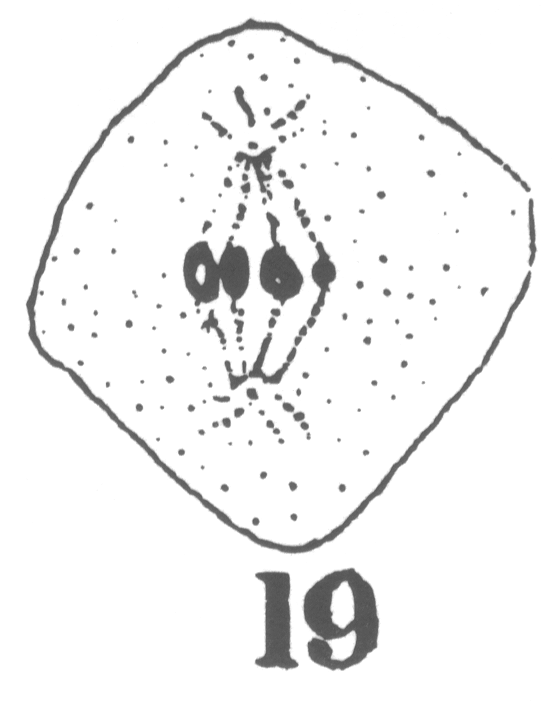
As the young spermatocyte grows, the sphere also increases in size, becoming more
and more rounded. From an early stage a minute centrosome (c)
is visible in the midst of the sphere substance. It is surrounded by a clear area, which
becomes more pronounced as the sphere grows older. Thus the developing cell gradually
acquires characteristics of size, shape and general appearance, that differ markedly from
those of the previous generation.
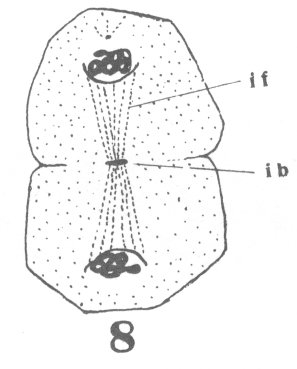
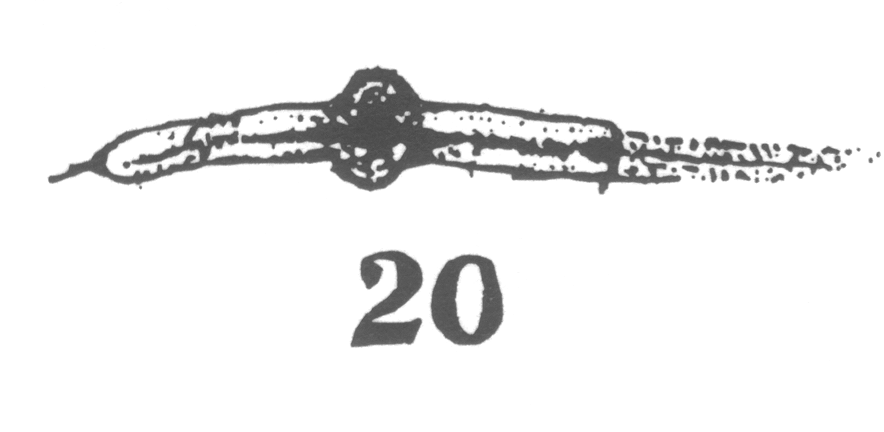
area closely applied to the nucleus, with
the horns of the crescent so extended as to
inclose more than half of the nuclear surface.
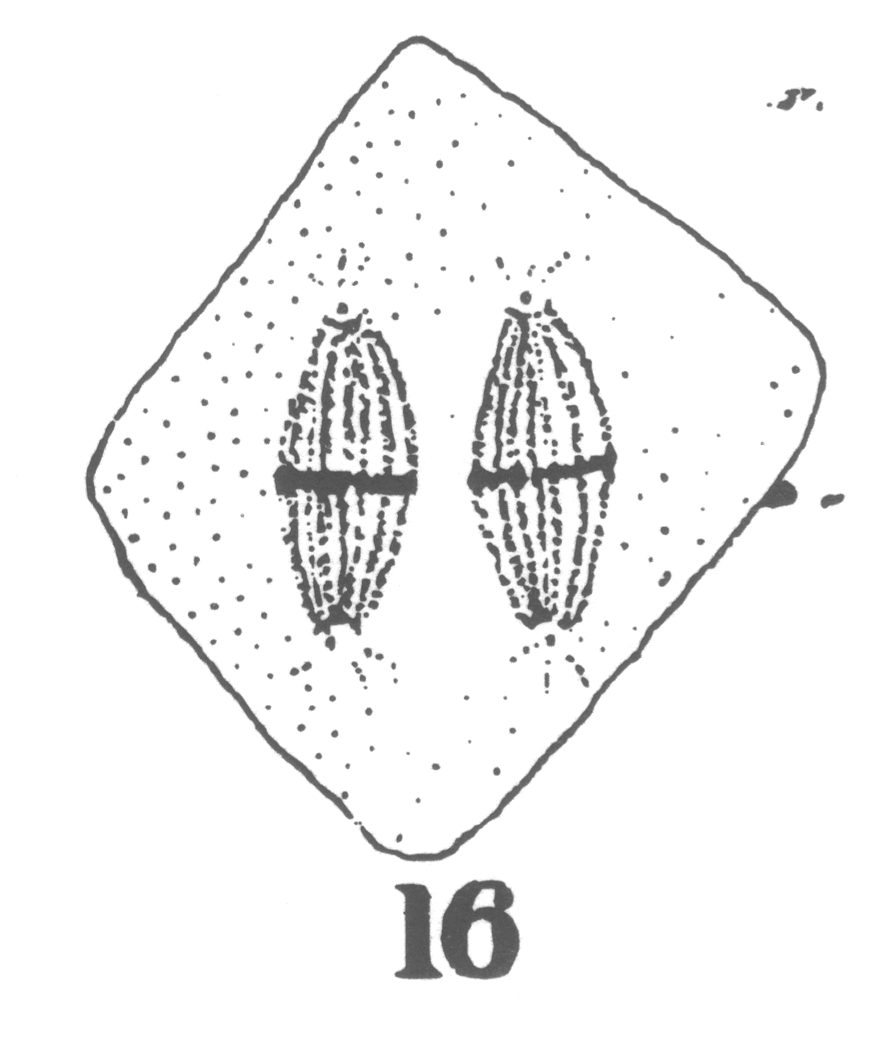
As the young spermatocyte grows, the sphere also increases in size, becoming more
and more rounded. From an early stage a minute centrosome (c)
is visible in the midst of the sphere substance. It is surrounded by a clear area, which
becomes more pronounced as the sphere grows older. Thus the developing cell gradually
acquires characteristics of size, shape and general appearance, that differ markedly from
those of the previous generation.