THE UNIVERSITY OF CHICAGO.
FOUNDED BY JOHN D. ROCKEFELLER.
Spermatogenesis of Normal and of Hybrid Pigeons.
A DISSERTATION
Submitted to the Faculties of the Graduate Schools of Arts,
Literature, and Science, in Candidacy for the
Degree of Doctor of Philosophy.
DEPARTMENT OF ZOOLOGY.
BY MICHAEL F. GUYER.
CHICAGO: 1900.
[Comments in square brackets, colour emphases and pigeon photographs have been introduced by DRF. Italicization is by MFG. Instead of being collected together at the end as in the original, drawings by MFG have been inserted into the text by DRF. Likewise, references originally presented as numbered footnotes have been inserted at the point of first citation.]
![]()
CONTENTS:
Introduction.
Methods.
General Arrangement of the Germinal Cells.A. Spermatogenesis of Normal Pigeons.
B. Spermatogenesis of Hybrid Pigeons.Spermatogonia.
Primary Spermatocytes:Early History.
Synapsis.
Division.Secondary Spermatocytes and their Divisions.
Reduction.Transformation of the Spermatid into the Adult Spermatozoon.
Abnormalities in the Structure of the Spermatozoon.
Degeneration of the Germinal Cells.
The Individual Histories of the Sterile Forms.
![]()
EXPLANATION OF PLATES I AND II.
[In the original text, Plate I contains Figures 1 -27 and Plate II contains Figures 28-50]
| a. | axial filament | s. | Sertoli or nurse cell |
| c. | centrosome | scy1 | primary spermatocyte |
| ch. | chromatin | scy2 | secondary spermatocyte |
| ec. | extra-nuclear chromatin | sg. | spermatogonia |
| h. | head | st. | spermatid |
| hs. | head-spine | t. | tail |
| i. | sphere or idiozome | tr. | tetrad chromosome |
| l. | nucleolar-like linin mass | v. | vacuole |
| o. | oil-globule | z. | intermediate body |
![]()
SPERMATOGENESIS OF NORMAL AND OF
HYBRID PIGEONS.
MICHAEL F. GUYER.
|
The present study on the spermatogenesis of normal pigeons is based largely on material obtained from two species, viz : Columba livia domestica, the common pigeon, Turtur risorius, the ring dove. Other species were studied for comparison. The particulars of the hybrid material will be given in detail in the latter part of this paper.
A description of the various phases undergone by the germ cells in their transformation from spermatogonia to mature spermatozoa is all that has been attempted in the present paper. It was intended originally to investigate also the origin of the spermatogonia and of the supporting or Sertoli cells, but it speedily became evident that such a study is a complicated problem in itself which must be postponed for the time being. On account of the comparative ease of manipulation and the abundance of material that can be secured at any time, the pigeon is in some ways well adapted to the study of spermatogenesis, but the small size of the cells makes a very high microscopic power necessary to obtain satisfactory results, and even then the cytoplasmic structure is insufficiently disclosed. So far, it seems that no investigations upon the spermatogenesis of the pigeon have been recorded. A few scattering observations have been made upon the spermatozoa, but the subject has been, in the main, untouched. Moreover, there has been practically nothing done toward studying the genesis of the germ cells in hybrids of any kind, although this field, from the very nature of the case, would seem to be an extremely promising one. The observations here recorded on hybrids are only the beginning of a number which I hope to make. To Professor C. O. Whitman I am indebted for much encouragement throughout the progress of my investigations. Had it not been for his generosity in supplying me with material, I could not have undertaken a study of hybrid pigeons at the present time. My thanks are also due to Doctor S. Watase for many valuable suggestions regarding methods. |
|
A number of different methods of fixing and staining were tried. Only the more important are mentioned. Flemming's strong solution was, taking all in all, perhaps the most satisfactory killing reagent. Nothing was found superior to it for faithful preservation of the cell, as a whole. It reveals the cytoplasmic structure better than other reagents, and at the same time brings out the chromosomes distinctly. Excellent results were obtained when it was followed by an aqueous solution of saffranin for twenty-four hours, and this by a counter-stain of lichtgrun in 95% alcohol. Immersion in the green for only an instant is required; if left for too long a time the saffranin is completely obscured. After using the green, the preparation must be washed rapidly in absolute alcohol, and plunged into clove oil. It may then be mounted in balsam either direct from the clove oil, or first transferred to xylol or cedar oil. Such a preparation shows the supporting or Sertoli cells to the best advantage, perhaps. The fatty globules present in them are not brought out so well by reagents which do not contain osmic acid. The green stain also brings the sphere substance very distinctly into view. Flemming followed by Heidenhain's iron-haematoxylin method also gave good results. Another killing fluid of almost equal value with the Flemming, is Hermann's Platino-aceto-osmic mixture. It is excellent when followed by a low per cent of alcoholic saffranin for 24 to 48 hours. After washing, the preparation is decolorized with acid alcohol and then treated with gentian violet, according to any of the well known methods. In the final clearing clove oil is used for a minute or two, and the preparation is then transferred to cedar oil or xylol. Too prolonged immersion in clove oil seems to extract the gentian violet. A reagent which is unsurpassed for the study of the individual chromosomes, the spindle and the centrosomes, is Gilson's mercuro-nitric fixing mixture, followed by iron-haematoxylin for staining. After such treatment, the chromosomes come out as clear cut as if stamped with a die. Tissues are cut into small pieces and left in the fluid from 4 to 6 hours. They are then washed thoroughly in 50% alcohol. Satisfactory preparations can be obtained only with thin sections measuring from 3 to 5 micra in thickness. To get the best results, the sections should be treated with the iron-alum for about 8 hours and then left for 36 hours in haematoxylin. After washing in water, they should be almost completely decolorized in the iron alum (15 to 30 min.). Thus, the haematoxylin is left in the chromatin only. The color of the haematoxylin may be deepened by a short immersion in a slightly alkaline solution before treatment with the plasma stains. Of the latter, either Bordeaux red or orange G. gives good results. To see the centrosomes to the best advantage the decolorization should not be carried so far. Among other valuable fluids should be mentioned that recommended by Lenhossek {1. Lenhossek, M. v: "Untersuchungen ueber Spermatogenese." Arch. f. mikr. Anat. LI, 1898.} in his paper on the spermatogenesis of the rat. The formula is as follows:Platinum chloride 1% = 50 cm. This gives a more faithful preservation of the cell cytoplasm and of the interstitial cells than Gilson does. It may be followed by almost any of the common stains. The iron-haematoxylin method gives very satisfactory results, as does also cyanin and erythrosin, or majenta followed by picric acid. This killing reagent has proved itself an excellent one also for general cytological work, especially in splanchnological research. For general purposes, according to Mr. A. D. Dunn, 1/4% platinic chloride may be substituted for the 1%, thus considerably reducing the cost of the fluid without materially interfering with its efficacy.
Graff's chrom-oxalic mixture was found valuable for both
chromatic and achromatic cellular structures. It was particularly serviceable in the study of the interstitial and wandering
cells. The formula which yielded the best results is the following: Tissues were left in this fluid from 2 to 4 hours, and then washed in 35% alcohol. Corrosive sublimate preparations, although badly distorted in many cases, were serviceable for comparison when followed by the Biondi-Ehrlich triple stain or by cyanin and erythrosin. The same may be said for picro-formalin-acetic preparations. The latter were invaluable in studying the centrosome, particularly during its transformation to form the axial fiber of the tail. Corrosive sublimate was substituted for the picric acid in some cases in the last named fluid, with the result that the staining of the axial filament was more precise. Besides the above methods, maceration was also employed and was of very great service in giving a general idea of the structure of the entire cell, the connection of the spermatozoa with the supporting cells, and the structure and transformation of spermatozoa. The most satisfactory results were obtained from Hertwig's well known macerating fluid. Various stains were employed after such treatment, prominent among which were acid carmine and methyl green.
In studying the transformation of the spermatid to the spermatozoon in sections,
the best results were obtained from
material fixed in corrosive sublimate and stained in Delafield's haematoxylin, with an after treatment of acid fuchsin and picric
acid. Both thick and thin sections were necessary. Gilson
material, followed by the iron-haematoxylin method, and counter-stained with Bordeaux red, was also good. |
GENERAL ARRANGEMENT OF THE GERMINAL
CELLS.
|
But little detail need be given regarding the general structure of the testis in the pigeon, for it does not differ essentially
from that of well known mammalian forms. The spermatogenesis of the rat is especially well known through the researches of Von
Ebner {1. Von
Ebner, V: "Zur Spermatogenese bei den Saugethieren." Arch. f. mikr. Anat.,
XXXI., 1888.},
Lenhossek {2. Loc. cit., p. 5.}
and others. In the pigeon
the testes are two elongated bean-shaped organs varying much
in size with age and with individuals. The germinal cells are
laid down in a very great number of convoluted tubules which
wind back and forth throughout the interior of each testis and
make up its main bulk. The tubules are much slenderer and more delicate than those of the rat, however, and can not be
isolated so readily for study.
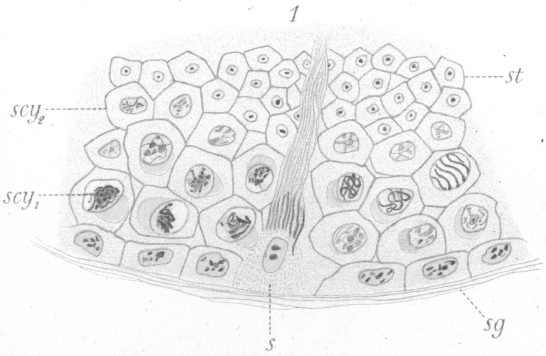
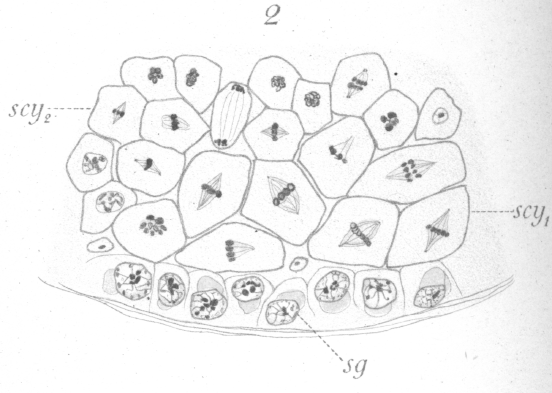
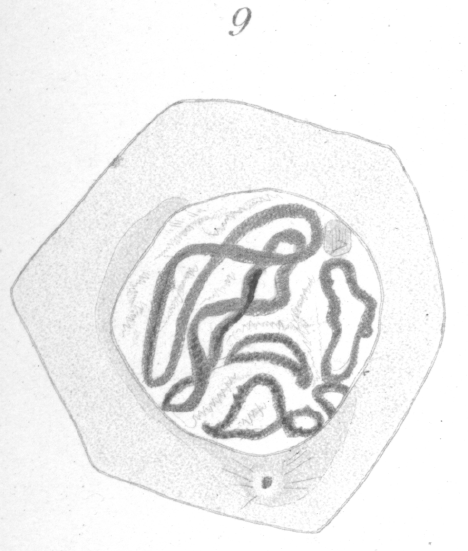
In the arrangement of the germ cells in the tubule, there is no departure from that of the other warm-blooded animals. Next the walls are the spermatogonia or parent cells (Fig. 1, sg.), which by growth and division give rise to the various generations of germ cells lying inward toward the lumen. The adult spermatozoa are formed through the final transformation of the spermatids, or cells produced by the last division, which lie nearest the center of the tubule. The ripe spermatozoa pass out through the lumen of the tubule and into the ducts which lead to the exterior. As in many forms, the spermatozoa attach themselves to a supporting cell (Fig. 1, s.) for a period before their complete maturation and ejection from the testis. The usual four phases or types of the germinal cells are recognizable, viz : (1) spermatogonia (Fig. 1, sg.), a more or less regular layer of cells lying next to the membrane or wall of the tubule, each cell of which through division gives rise to two new cells. One or both of these may increase in size and become (2) primary spermatocytes (Fig.1, scy1), or remain in the layer and continue as spermatogonia. The primary spermatocytes, after some interesting changes, divide to form, (3) the secondary spermatocytes (Fig. 1, scy2), which divide again shortly to give rise to (4) the spermatids, through the transformation of which the spermatozoa are developed. The number of chromosomes in each type, as seen at the equator of the spindle before division is, in the spermatogonia 16 loops, in primary spermatocytes 8 rings or vesicles, and in secondary spermatocytes 4 rings or vesicles. It is not improbable that the socalled ring chromosomes, at least under certain conditions, are really small vesicles bearing the chromatin in the form of an internal shell. To decide this matter has proved the most tantalizing point in the whole investigation. Such a vesicle could easily take on the appearance of a ring when viewed by transmitted light and from the fact that from any point of view many of the chromosomes present in a given field appear to be rings one is inclined to accept this idea of their vesicular nature. On the other hand the chromosomes, of the primary spermatocytes at least, undoubtedly originate as rings although it is possible for them to become vesicles as they condense. What appears to be the elliptical edge of a thick ring is presented occasionally, and this has decided the writer to use the term ring in preference to vesicle, although it should be borne in mind that the apparent rings may be vesicles. Closely connected with the germinal cells proper are the supporting cells or Sertoli cells mentioned above, to which the spermatozoa become attached at one period of their transformation (Fig. 1, s.). They are irregularly disposed among the other cells and frequently do not lie in the spermatogonial- layer. It is difficult to get preparations showing only certain typical stages, for while certain kinds of cells may be present in one view, in another, some of exactly the same phases may appear side by side with other cells in entirely different stages of development. There seems indeed to be no very definite sequence of stages in a given part of a tubule as we find, for instance, in the rat. To be more specific, one may find spermatogonia, Sertoli cells with spermatozoa attached, and spermatids in one view. In another, where the spermatogonia are apparently in the same condition as before, primary and secondary spermatocytes may be present. Where one finds the primary spermatocytes, they are usually arranged in clumps, the individuals of which are at similar points of development. The same is true for the secondary spermatocytes. Fig. 2 shows both kinds of spermatocytes undergoing division in the same field of view.
In the following account, the spermatogonia are considered first and the various other types are then taken up in the order of their succession in development into spermatozoa. |
A. SPERMATOGENESIS OF NORMAL PIGEONS.
|
The spermatogonia lie in a more or less regular layer next
to the tubule wall. Not infrequently the individual cells have
the appearance of having been crowded out of place so that the
layer appears irregular or double. There is considerable variation in the appearance and some variation in the size
of the
cells at different times. In early stages they are far apart and
have small nuclei which are oblong with the long axis parallel
to the tubule wall. The cells themselves are generally flattened
more or less so that their long axes coincide with those of the
nuclei (Fig. 1, sg.). In still earlier stages, the cell boundaries
are very indistinct or seemingly absent, and gaps frequently intervene between the individual
spermatogonia, so that they appear to have been left behind from a preceding set, or
to have recently settled in their present position. Some of them resemble very much the wandering cells seen outside the tubule.
In later stages, the spermatogonia are crowded together
until they become more or less columnar in shape (Fig. 2).
The nuclei increase in size and become very distinct. They are
round or slightly oval in shape. When oval, the long axis is,
as a rule, perpendicular to the tubule wall. In the closely
packed cells, the nuclei come to lie eccentrically as they enlarge,
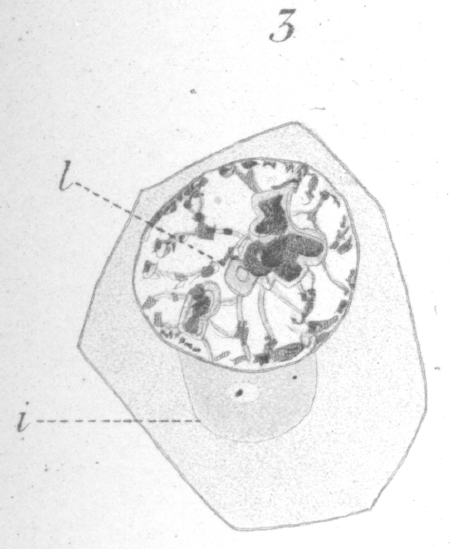
and a condensation of the cytoplasm, or mass of sphere substance (idiozome of
Meves), makes its appearance and
gradually increases in size till it becomes a well defined area (Fig.
3, i). At first it seems to be entirely granular in nature, but
later displays, at least in part, a fibrous or reticular structure.
Near the center of the mass is generally a clear area in which
a minute centrosome is discernible. As the sphere itself becomes better defined, the centrosome appears larger and the
clear area more distinct (Fig. 3). Fig.
3. x 2000 -
A spermatogonium in the resting condition.
A sphere is often seen in the young spermatogonia just before division when they are still lying far apart; but, it, as well
as the nucleus, seems to differ from the sphere and nucleus
that make their appearance just before the last spermatogonial
division, subsequent to which the resulting cells undergo a
period of growth to become the primary spermatocytes. During this last nascent period, the sphere is very large in proportion to the cytoplasm and lies typically on the side of the nucleus
next the tubule wall. Not only the cells which have divided off
towards the lumen of the tubule form spermatocytes; those
which have remained next the wall may double out toward the
lumen of the tubule, increase in size, and also develop into
primary spermatocytes.
Rarely they remain in place; in such
an event, one finds the primary spermatocytes and ultimately
the spermatids, next the wall of the tubule.
In the resting condition of the spermatogonial nucleus, the
chromatin is visible ordinarily as small, deeply stained granules
distributed along the nuclear membrane, and upon the loose
network which extends throughout (Fig. 3). The granules
along the periphery are the smallest. There are usually from
one to three or four chromatin masses or chromatin nucleoli of
varying size, present at or near the center of the nucleus.
There is frequently discernible a distinct clump of material
in the nucleus which reacts to stains in the same way as the
linin and which may perhaps be considered as an achromatic
nucleolus. It seems to be in direct continuity with, in fact a
part of, the linin (Fig. 3, 1.) McGregor
{1. McGregor, H: "The Spermatogenesis of
Amphiuma."-Journ. Morph. XV., Suppl., 1899.} has made a similar observation
in the spermatogonia of Amphiuma. In the
pigeon, when the saffranin gentian-violet method is used, both
the linin and the nucleolar-like mass take the same bluish violet
color; likewise, both react similarly to Bordeaux red. The mass may persist as a distinct body in the
nucleus during the
preparatory rearrangement of the chromatin for division (Fig.
4, 1.), but it usually takes part, together with the network, in
forming the achromatic sheath or covering within which each
individual chromosome is incased.
Before division the nucleus passes from the so-called resting
condition through an incomplete spirem stage. The spirem is
formed through the rearrangement of the chromatin and linin
material. The chromatin is laid down in minute granules, and
these begin to spread out from the various chromatic dots and
clumps through the linin substance which forms a sort of covering or case for them. If the saffranin gentian violet method
has been employed in staining, the linin is of a reddish violet
color and the chromatin granules deep red. As the granules
extend out more and more along the violet threads, the latter
become redder and redder, until in the final condition of independent chromosomes, each chromosome appears as a red loop
or thread. A very close examination under high power reveals
the fact, however, that the red part of the chromosome is surrounded by a thin violet colored layer of linin
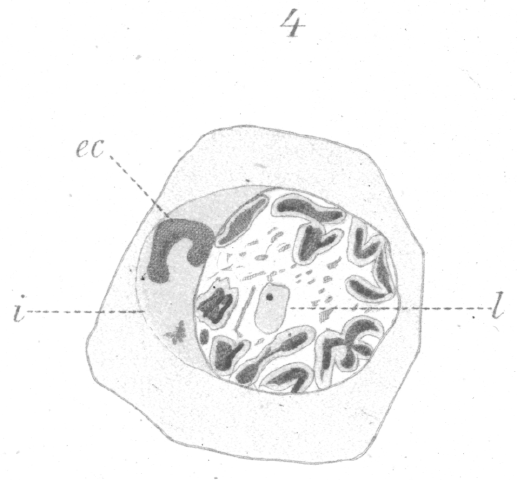
(Fig. 4).
At first the newly formed chromosomes lie scattered irregularly throughout the nucleus, but later, just before the spindle
is formed and the nuclear membrane disappears, they arrange
themselves around the periphery. Sometimes the nucleolus of linin-like material persists and is visible lying more towards the
center of the cell (Fig. 4, 1.). With the transformation of the
spirem into chromosomes there is also an increase in the volume of the nucleus.
In the last division of the spermatogonia before they undergo growth to form spermatocytes there is a curious casting
out of part of the chromatin into the cytoplasm, while the
chromosomes are yet in an early stage of formation. After the
spirem breaks up into individual chromosomes, the latter are
visible as irregular threads and loops scattered throughout the
nucleus. They appear later as small rods or loops which are,
perhaps, elongated vesicles with an internal shell of chromatin,
judging from the fact that they frequently have the appearance
of a lengthened ring. Amongst this colony of loops and rods
there is always one chromatic mass that differs very much from
the others in size. It is a filament two or
three times as long
as the other chromosomes and it never has the ring or split
appearance. Before the final movement of the chromosomes
to the periphery of the nucleus, this long filament is thrown out
into the cytoplasm (Fig. 4, ec.). In all cases examined it was
cast out into the sphere substance which contained the centrosome and lay close to one side of the nucleus.
The body thus ejected breaks up apparently, and becomes
scattered throughout the cytoplasm. The significance of this
phenomenon could not be determined. Whether the mass is
simply a useless accumulation of chromatin, or whether it is
of more vital importance to the phenomenon of maturation
seems beyond explanation at present. There is apparently no diminution in the
number of chromosomes, yet from its reactions to stains the ejected filament
seems to be purely chromatic in nature. The chromosomes are so massed in the
spermatogonia, however, that a mistake in the count might easily be
made. There were indications that in some cases the staining
of the filament in question was a trifle deeper than that of the
other chromatic elements, but the difference was ordinarily
scarcely perceptible. Fig. 4 shows one of three adjacent cells
in all of which the process was in progress.
It is questionable if the extruded body can be homologized
with the "accessory chromosome" of McClung
{1. McClung, E. C.
A Peculiar Nuclear Element in the Male Reproductive
Cells of Insect: Zool. Bull, II, 4, 1899.}, a curious
nuclear element which he describes as occurring in
Xiphidium
fasciatum,
one of the
Locustidae,
and which he thinks is perhaps
identical with the chromatin nucleolus described by Wilcox
{2. Wilcox, D. V.
Spermatogenesis of Calloptemus femur rubrum and Cicada tibicen: Bull of the Mus. of Comp.
Zool., Harvard College XXVII, 1.}
in
the testicular cells of
Caloptemus femur rubrum,
or in Pyrrochoris
as recorded by Henking
{3.
Henking, H. Erste Entwickelungsvorgange in den Eiern der Insecten.
Zeitschr. f. Wiss. Zool. LI. 1891.}. Again in Montgomery's
{4. Montgomery, Th. H. The Spermatogenesis of
Pentatoma: Zool. Jahr.
XII, 1898.} account of
Pentatoma,
McClung finds a possible parallel in a body which
resembles a nucleolus while the cell is at rest but simulates a
chromosome during the period of division. There is no evidence in the case of the pigeon to show that the ejected
filament is derived directly from a chromatin nucleolus. The
chromatin nucleoli when present lose their identity apparently
and are absorbed or built up into the spirem when the preparations for division begin. Moreover, the accessory chromosome
of McClung
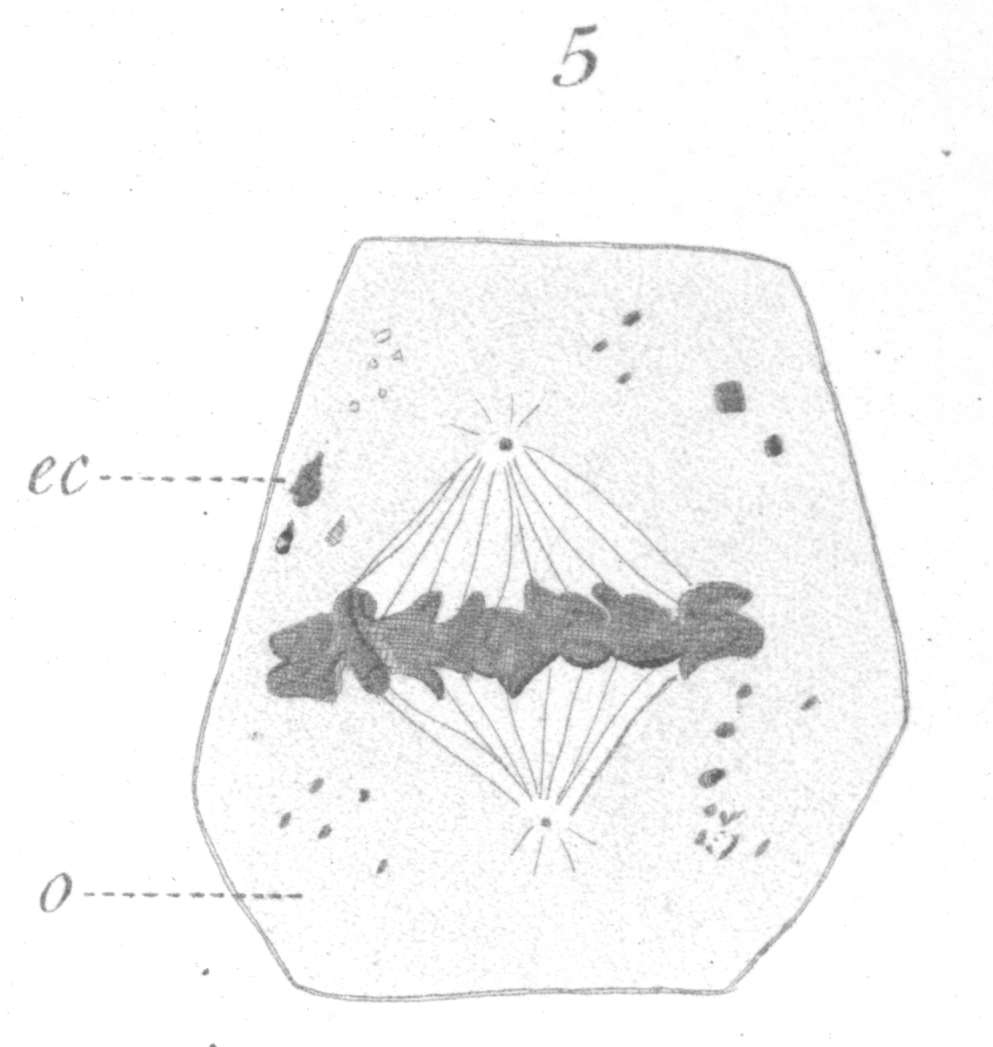
Fig. 5. x 2000 - A spermatogonium ready for division.
After the chromosomes have undergone their transformation inside the nucleus and have approached the nuclear membrane, the centrosome which is seen lying in a clear area of the
sphere, divides into two, one of which moves along the outer
periphery of the nucleus to, the opposite pole. The first appearance of the spindle fibers is as radiations which spread
around the nucleus from the centrosome. No further detail
could be made out regarding the origin of the spindle. The nuclear membrane which has persisted in a very
distinct form so far disappears and the next stages show the chromosomes in connection with the spindle. The polar radiations extend only a short distance until lost to view. They
are but poorly developed at any of the divisions. Fig. 5 shows a side view of
spermatogonium just ready for division. The spindle
is short and broad and the chromosomes lie in a confused band at the equator.
The individual chromosomes are for the most part in the shape of short bent rods or loops, with
the closed end of the loop turned towards the center of the spindle.
Their form, which can be recognized only with difficulty, is seen to best
advantage in a favorable view from one pole. They often split longitudinally before the spindle is formed
and this adds to the confusion when massed. In dividing, if they have not already halved before coming to the
spindle, they split lengthwise, beginning at the inner closed end. While moving towards the poles it is
not uncommon for the free ends of a chromosome to fuse, thus forming a small ring or
globule. In some cases a few of the loops may be in the form
of apparent rings even at the equator. The distance between
the centrosomes increases somewhat as the division proceeds.
In the division of the spermatogonia, the general appearance of the spindle and the arrangement of the chromatin in
loops seems to be much the same as that of the ordinary tissue
cells of the pigeon as exemplified in leucocytes and the cells of
the liver, adrenal and kidney. The chromosomes are crowded
together so closely and the loops so irregularly disposed as to
render an accurate count very difficult. Repeated attempts at
counting both before and after the equatorial plate stage point
to sixteen as the correct number.
After division the mass of chromatin becomes so fused together and dense in appearance as to prevent any determination of the details of nuclear reconstruction. At each pole a
clear area is maintained between the centrosome and the
chromatin for some time while the nuclear membrane is forming on the side of the latter which faces the equator but the
centrosome is finally lost to view and can not be seen again.
When a centrosome appears later in the sphere substance of the
spermatocyte it seems to be generated anew.
A system of connecting fibers extends from one nucleus to
the other for a time; as preparations for complete cell-division
advance they converge towards the equator more and more.
A distinct intermediate body or "zwischenkorper" is visible,
marking out the path of the approaching cytoplamic division.
The persisting threads often show small varicosities or granules which react to reagents like the centrosome
and mid-body.
During the nuclear division the cytoplasm has the appearance
of a homogeneous granular substance with oil globules and the
remnants of the extruded chromatin scattered about irregularly
through it (Fig. 5). Shortly after the cell-division is completed
these deeply staining granules have all disappeared. It seems
probable that they have been consumed in some way by the
cytoplasm which increases rapidly in bulk.
|
|
The primary spermatocytes or cells of the second spermatogenetic period originate, as has been indicated, from the
cell-products of the last spermatogonial division through a
process of growth. All transitional sizes between that of the
spermatogonia and the comparatively large spermatocytes may
be seen in practically the same order as that described by Lenhossek {Loc. cit., p. 5.}
for the rat. In the new cells, the chromatin passes into the resting condition and an increase in bulk of both
nucleus and the cytoplasm begins. The chromatin is arranged
much as it was in the resting condition of the spermatogonia
except that there is perhaps less distribution of the chromatin
granules along the periphery of the nucleus and the linin fibres
are coarser (Fig. 6).
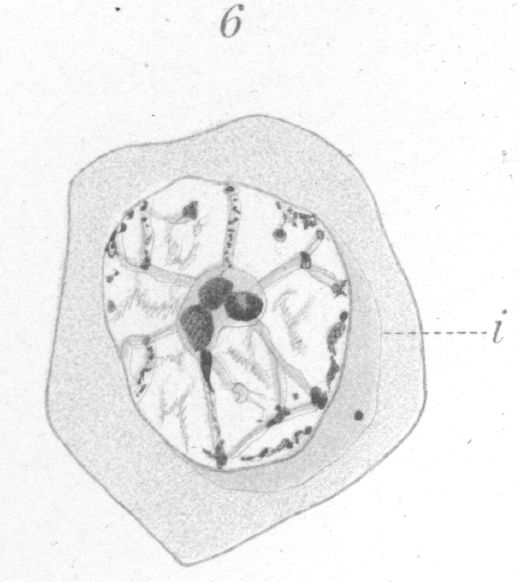
The sphere first appears as an indistinct granular crescentic area closely applied to the nucleus, with the horns of the crescent so extended as to enclose more than half of the nuclear surface (Fig. 6, i.). As the young spermatocyte grows, the sphere also increases in size and become more and more rounded. From an early stage a minute centrosome is visible in the midst of the sphere substance. It is surrounded by a clear looking area which becomes more pronounced as the sphere grows older. Thus the developing cell gradually acquires characteristics of size, shape, and general appearance that differ considerably from those of the previous generations. At no point in this metamorphosis could I distinguish any nuclear element that might be identified positively as an achromatic nucleolus. Such a body, in fact, seems to be wanting in all stages of the spermatogenesis. One or more bodies of varying size were often present but there was no evidence whatever to show that they differed in any way from the ordinary linin material. Certainly no such well marked object exists as Moore {1. Moore, J. E. S. On the Structural Changes in the Reproductive Cells During the Spermatogensis of Blasmobranchs. Quart. Journ., XXXVIII, 1893.} figures in the testicular cells of the elasmobranchs, or Lenhossek {2. Loc. cit., p. 5.} in those of the rat.In the growing cell the cytoplasm has the appearance of a granular mass throughout which fine ramifying fibres can be detected occasionally. The sphere at times exhibits a delicate fibrous structure but this was not observed frequently and may have been due simply to the action of the reagents. |
|
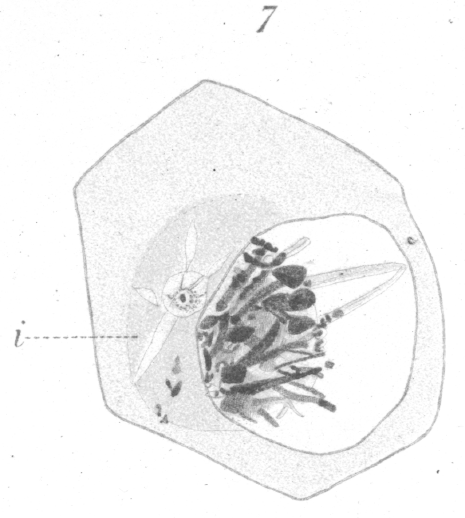
As soon as the growth of the primary spermatocyte is completed, the nucleus, which is voluminous and apparently turgid
with sap, undergoes a curious change. There is a marked
drifting of the nuclear contents to that side in contact with the
sphere (Fig. 7).
This phenomenon is the same that Moore recognized in the maturation of Elasmobranch sperm cells and
which together with the accompanying fusion of pairs of chromosomes he has designated by the term
synapsis. A similar
union of the chromosomes occurs in the, pigeon, for at the ensuing division only
eight chromosomes appear. Thus, a
pseudo-reduction in Ruckert's {3.
Ruckert, J: Zur Eireifung bei Copepoden: An. Hefte, 1894.}
sense occurs.
In the collapsed and tangled mass of the nuclear elements that have passed to one side of the nucleus, no distinct arrangement can be recognized. The remainder of the nucleus seems to be absolutely devoid of any contents which will react to stains. Often one or more stiff bristle-like fibres or strands of what appears to be the linin material project out from the mass across the vacant area of the nucleus. The ends of shorter fibres are seen sometimes bristling out from all sides of the heap. If saffranin and gentian violet are used for staining, one observes granules and clumps of chromatin taking a deep red stain, mixed indistinguishably with a bluish or violet mass of linin. No distinct chromosomes as observed by Montgotnery {4. Loc. cit., p. 12.} in Pentatoma could be seen. Chromosomes could be distinguished in fact, only after the nucleus had passed through a spirem stage, when, as has been noted already, the chromatin appears laid down in half the original number of chromosomes; that is, the new chromosomes are of the bivalent type. As to the meaning of synapsis, we seem to be in almost total ignorance. Few suggestions seem to have been proposed to explain it beyond that of Moore {. 1. Loc. Cit., P. 16.} who mentions the possibility that it may mark an abortive attempt to bring about the formation of a tailed spermatozoon, due to a sort of historical reminiscence which marks this as a once final stage of development in the remote ancestral sex cells. It seems to the writer more probable that the fusion is a conjugation of maternal and paternal chromosomes, though why such is necessary, is not apparent. Some rather significant facts appear which may furnish a possible clew to the nature of some of the changes which occur during the period of nuclear collapse. A very careful determination of the size and shape of the nucleus and sphere substance respectively was made in a number of cases just before and during the contraction of the contents to one side. Immediately before the change, the large nucleus is nearly spherical, or if oval, its long axis may lie in any direction with reference to the sphere. After the contraction has occurred, the nucleus is always slightly flattened toward the sphere; that is, the long axis of the oval lies parallel to the sphere. Moreover, at times the nuclear membrane is a trifle wavy or uneven. The average diameter of the nucleus before is slightly greater than during the period of apparent collapse. A large number of measurements of the sphere reveals the fact that it is, on an average, almost a third larger after the chromatin has massed nearer it than before. This, together with the fact that the drifting of the nuclear contents is always toward the sphere, and that the nuclear membrane is indistinct in that region, leads to the conclusion that there has been a discharge of part of the nuclear material into the sphere. Then, too, occasional fragments of chromatin are seen scattered about in the sphere, indicating that they have been carried out, perhaps during a discharge of the material from inside the nucleus. Again, the sphere takes stain more intensely and appears more dense as soon as the above change has occurred and there are visible frequently minute star-like radiations of a lighter staining substance which spring out from the clear area around the centrosome (Fig. 7, i.). The radiations remind one of the clear-looking streaks which accompany the formation of the pulsating vaculole in some of the infuseria. What the interpretation of the above facts is, can be answered at present only conjecturally. The first thought that presented itself was that the centrosome originally lay inside the nucleus and that the above occurrence was a process of extrusion by which it reached the sphere. But careful examination of the cells reveals the presence of the centrosome in the sphere long before synapsis. Indeed it seems to originate from the sphere substance itself and to increase in size as the, sphere develops. It is possible that the passage of substance from the nucleus to the sphere may have something to do with the formation of the spindle, for immediately after synapsis the centrosome divides and the spindle fibres appear. It may be that some substance is required from the nucleus before the extremely heavy spindle of this division period can be constructed. The sphere in the germinal cells of the pigeon seems to be simply a very plastic area of the cytoplasm especially favorable for building up into such structure as spindles, asters, centrosomes, the apex of the head, and the axial fibre of the spermatozoon tail. That such a role on the part of the sphere is of very general occurrence in germ cells is so well known to workers in spermatogenesis as not to require specific reference. In the ovarian egg the sphere (idiozome) seems to be employed likewise in diverse ways. In one case of abnormal ovarian structure in the pigeon (Guyer {1. Guyer, M. F: Ovarian structure in an Abnormal Pigeon.--Zool. Bull. II. No. 5, 1899.}) where numerous multiple eggs were forming and extensive degeneration going on, the sphere seemed actively employed in building up cell membranes as well as in the formation of fibres. The large vacuoles which were present in many eggs, moreover, originated invariably in the center of the sphere. In other words, the sphere is, then, only an active condition of the cytoplasm in a given region. The greatest visible change is in the nucleus itself at this period of disturbance in the primary spermatocyte. The chromatin drifts to one side and during its stay must undergo some sort of a profound alteration in its arrangement, for when the chromosomes reappear they are the double or bivalent type. It is possible, however, that we emphasize this phase of the phenomenon simply because it is the more visible through the great staining capacity of the chromatin. That the pseudoreduction may occur in reproduction cells without the accompanying phenomenon of intra-nuclear collapse is well seen in the primary spermatocytes of the rat, where such a condensation seems never to occur, although only half the original number of chromosomes appears at the time of division. In one or two instances it seemed very probable that during the internal changes of the nucleus quite enough chromatin to form a complete chromosome or more was ejected into the cytoplasm. At least two distinct examples were seen where there was a very tiny spindle connected with chromatin material lying in the cytoplasm at considerable distance from the main spindle. In one case, there was a single chromosome attached to the dwarf spindle, and in a second there was one large and one very small chromosome. Unfortunately the number of chromosomes at the equator of the main spindle could not be determined in these instances. It is interesting to note in this connection that the chromatin apparently determines the development of the spindle, for where we have enough chromatin accumulated to constitute a complete chromosome, it would seem that a spindle, develops. Juel {1. Juel, H. O: Die Kerntheilungen in den Pollemutterzellen.-Jahrb. wiss. Bot., XXX, 1897.} in a recent paper records an apparently similar occurence in the pollen-mother-cell of Hemerocallis. On the other hand, the possibility exists, of course, that the small spindle may have been simply a readjustment of a part of the main spindle together with a chromosome which had become separted from it in some manner. |
The Division o f the Primary Spermatocyte.
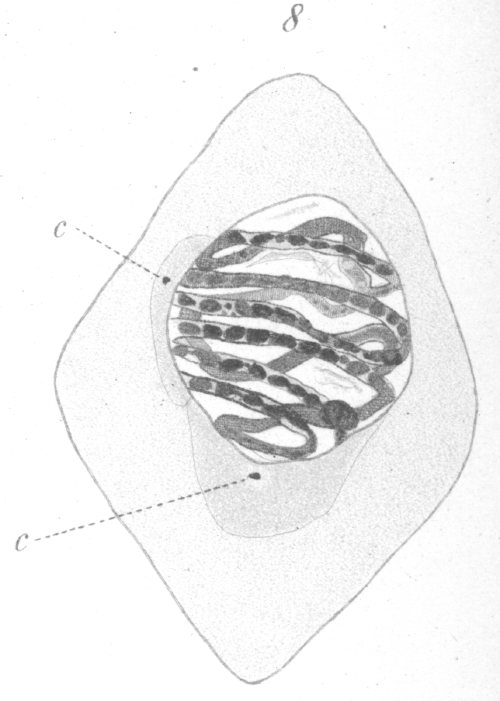
As soon as the phase of synapsis is past, the nucleus undergoes a process of reconstruction and the contents are once more redistributed throughout the whole internal area. It does not pass back into a resting condition, but proceeds at once to the development of the spirem. In reconstruction, the linin fibers are the first to spring out from the eccentric mass and gradually extend throughout the nucleus. The chromatin soon follows until a large well developed spirem is formed (Fig. 8). As in the spermatogonia, the chromatin substance seems to be laid down in the form of granules which are imbedded in the linin fibres. The granules spread until they are finally about equally distributed throughout the linin. In the early stages of spirem formation, chromatin nucleoli or net knots are present, but they gradually resolve into granules which spread until all portions of the spirem are approximately equalized in the supply of chromatin. Looked at from certain views, the spirem seems to be a continuous thread, but from others it appears as a number of closed loops. Whether these loops exist as such from the first could not be determined.
Early in the reconstruction after synapsis, a division of the centrosome occurs and one of the new centrosomes move slowly along the outside of the nuclear membrane toward its place at the opposite pole of the nucleus. As this centrosome advances, a part of the sphere substance spreads around the nucleus and accompanies it (Fig. 8). The nuclear wall persists as a distinct membrane till late in the preparation for division. Concomitantly with the migration of the centrosome, the spirem-like appearance inside the nucleus disappears gradually, until by the time the centrosome reaches its position at the opposite pole of the nucleus, the components of the spirem are seen distributed through the nucleus in the form of 8 elongated, irregularly-shaped rings (Fig. 9). If the saffranin-gentian-violet method of staining is used, the rings can easily be seen under high power to consist of a violet groundwork of linin in which are imbedded numerous red granules and lumps of chromatin, so that the strands have a monilated appearance. Occasionally one or two violet staining nucleolar-like masses of linin exist independently in the nucleus (Fig. 9).
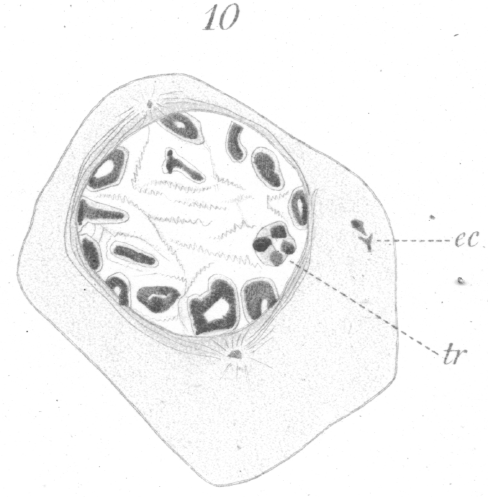
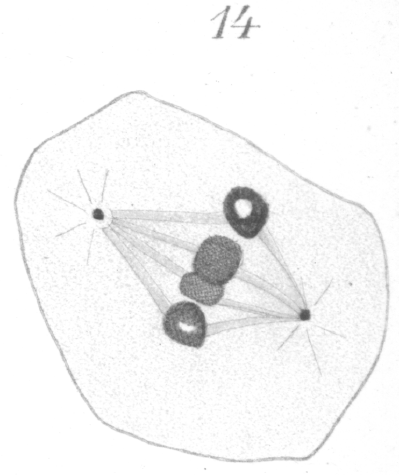
Further preparation for division is seen in the gradual condensation of the elongated rings into a shorter and heavier type. The chromatin becomes fused together gradually in such a way that the distinct granules are no longer visible. It is not unusual, after the condensation has proceeded for some time, to see some of the rings in the form of a four-partite structure as shown in Fig. 10, tr., where in one of the chromosomes to the right, four more or less spherical densely staining areas connected by lighter bands of linin are visible. This is by no means universal. It is possibly to be compared to the tetrad formation so frequently observed in maturation phenomena. Such a chromosome is strikingly similar in appearance to the tetrad types pictured by vom Rath {1. vom Rath, O: Zur Kenntniss der Spermatogenese von Gryllotalpa vulgaris.-Arch. f. mikr. Anat., XL, 1892.} for Gryllotalpa, or Wilcox {2. Loc. cit., p. 12.} for Caloptemus. It might be well to mention here, also, that in the primary spermatocytes of hybrids in some instances, one or more of the large chromosomes was observed to consist of four small closely apposed rings or vesicles (Fig. 35).
When the final condition of chromosome formation is attained, the chromosomes exist as eight thick rings. Examination under high power shows that each ring consists of a thread or shell of chromatin encapsuled in a film of linin. If the preparation is not sufficiently decolorized after staining with haematoxylin, the chromosome will have the appearance of a small black disc or globule, its ring-like nature being entirely obscured. The chromosomes, after completing their development, gradually arrange themselves along the thick nuclear wall (Fig. 10), as in the corresponding stage preparatory to division in the spermatogonia. A residue or condensation of linin material forming a nucleolar-like mass may be present near the center of the nucleus.
By the time the nucleus has assumed this arrangement, the
fibers of the spindle have become much more distinct and
spread around the nucleus from either pole toward the equator (Fig. 10). Small astral radiations are also visible, but they are
very scant and can be traced only a short distance from the centrosome.
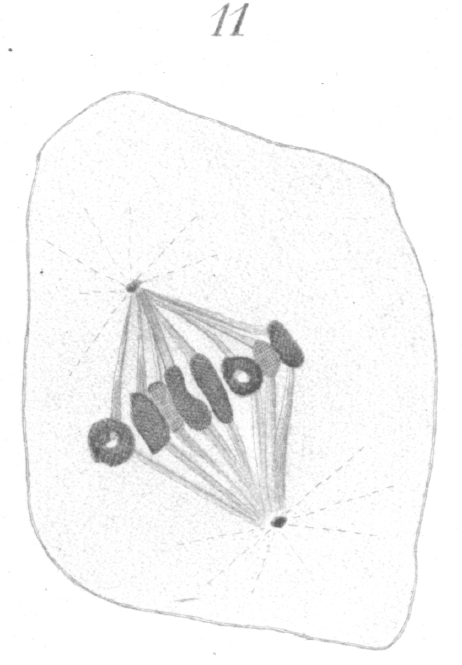
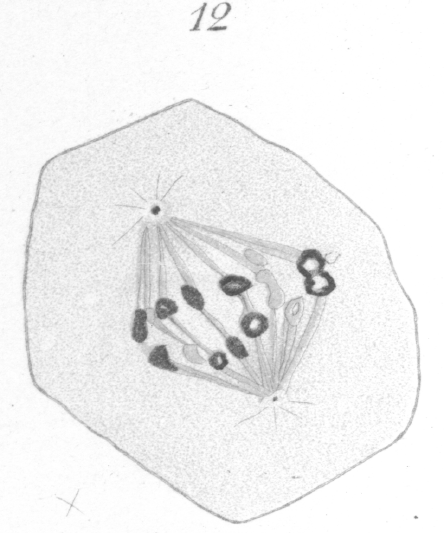
The chromosomes stand out prominently from the spindle at first, as if attached by only one edge, but they finally become more closely affixed. Considerable practice is required before any satisfactory results can be obtained in counting. Inasmuch as the chromosomes do not lie in one plane, careful manipulation of the fine adjustment of the microscope is necessary and a large number of cells must be examined. Even then the results are not always as conclusive as one might wish. In the present case, nevertheless, I think there is little doubt that eight is the correct count. In dividing, the chromosomes always elongate toward the poles and break transversely at the equator. They may stretch to a considerable distance, before the final rupture occurs. Often the chromatin part of the ring divides early and the two daughter chromosomes remain connected by a thick band of the achromatic substance which encases them (Fig. 12). These connecting strands or tubes of achromatic material, indeed, persist generally until the chromosomes have extended quite to the two poles. As soon as the division of the chromatin occurs, the free ends of each of the newly formed chromatic loops almost invariably fuse together immediately to form a ring or vesicle which is smaller and thinner than the original mother chromosome (Fig. 12).
The large thick spindle fibers do not seem to persist at the equator after the chromosomes have divided. They may be seen extending from the chromosomes to the pole as long as the chromosomes are moving poleward, but when the chromosomes have reached their goal, the fibers disappear. A system of distinct interzonal fibers exists, but they are plainly the linin filaments which have been left stretched between the separating chromosomes, and are not the remnant of the original spindle. They are much more delicate than the spindle fibers and have an entirely different appearance. Occasionally bits of chromatin may be seen imbedded in their substance (Fig. 13). As the division proceeds these fibers gradually constrict in the region of the equator and acquire the well known hour-glass shape. They ultimately fuse at the equator and form a distinct band-like intermediate body (Fig. 13) which may persist even after the new cell walls are formed and the cells are no longer contiguous.
Before the final division of the cytoplasm is completed, the nuclear reconstruction is well under way. A clear area between the centrosome and the chromosomes persists for some time, but it finally becomes obscured and the centrosome is lost to view. The new nuclear membrane forms first on the side of the nucleus away from the centrosome. At times the chromosomes break up into a number of smaller bodies or granules while yet in transit toward the pole, but usually they remain intact. As soon as the chromosome cease their progressive motion, and sometimes before, they fuse with one another (Fig. 13) into a dense mass which ultimately spreads about in such a way as to form the new network with its distribution of chromatin. None of the details of this process could be determined because, after the vesicular condition consequent upon the fusion of the rings, very few stages could be found. This fact would indicate that the process is one of very short duration. It is doubtful in some cases, indeed, if they go into a resting condition before the next division.
While the above transformations are occurring in the nucleus, the cytoplasm seems to remain in a quiescent state. It
has the general appearance of a homogeneous substance in which at times a delicate network is visible. Occasional particles of chromatin may be scattered about in it (Figs. 10, 13). The product of the division just discussed consists of two cells which are of considerably smaller size than the primary spermatocyte itself and which never attain to its volume. Their nuclei, too, are smaller than the parent nucleus and compare more nearly in size with those of the spermatogonia. These cells are the spermatocytes of the second order, or secondary spermatocytes. A distinct resting stage, occurs in some of the secondary spermatocytes at least, but this may not be true for all. It is a very transient condition, however, and the cell proceeds almost immediately to division. In the preparation for division there is nothing which corresponds to the collapse of the nuclear material in the primary spermatocyte, nor is there formed at any time a well marked spirem. The chromatin seems to pass rapidly from the apparent resting condition into one where it exists as distinct chromosomes. It is very easy to find numerous cells in process of division, but the stages preparatory to division are rarely seen. This would lead to the conclusion that perhaps in some cases the chromosomes rearranged themselves for the second division without passing through a resting stage. When the chromatin takes on the form of chromosomes, curiously enough only four rings or vesicles appear instead of eight, as seen in the primary spermatocytes. This number persists and as a result only four chromosomes come to the equator when the spindle is formed (Fig. 14). Occasionally a spindle bearing eight chromosomes of a smaller size is to be found. The writer was unable to decide whether such a type belongs also to the secondary spermatocyte or whether reagents have simply caused an unusual shrinkage in the chromosomes of the primary spermatocyte.
The formation of the spindle and the attachment of the
chromosomes to it occurs in much the same manner as in the
primary spermatocytes and will require no detailed discussion.
The final size and appearance of the individual chromosomes is
practically the same as that of the bivalent chromosomes of the previous generation, although only half the number is
present. There may exist also a residual nucleolar-like mass
of linin at some point in the nucleus. The spindle is a trifle
shorter and slenderer than that of the previous division. The
number of fibers appears to be four, thus again corresponding to with the number of chromosomes.
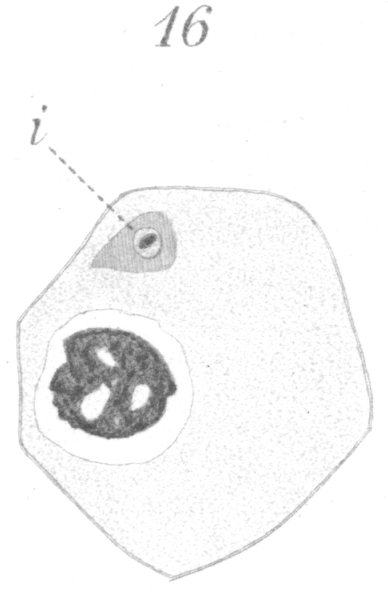
The reconstruction of the daughter nuclei differs in some respects from that of the preceding generation. The chromosomes do not fuse so readily, but lie, side by side for some time in the form of four hollow vesicles. These usually fuse later with the result that one large hollow sphere of chromatin is formed near the center of the nucleus, giving the spermatid a very characteristic appearance (Fig. 1, st.). Numerous fine fragments of chromatin migrate to the nuclear membrane which has appeared in the meantime, and form a thin shell along its inner surface. Fibers of linin radiate out from the central mass to the periphery of the nucleus. The centrosome persists and together with the tip of the spindle moves out into the cytoplasm to one side of the nucleus after the nuclear membrane is formed (Fig. 16, i.). The remnant of the spindle which surrounds the centrosome seemingly becomes reconverted into sphere substance. The new cell thus formed from the division of the secondary spermatocyte is the spermatid and is the cell which will ultimately be transformed into the spermatozoon.
In the division of the spermatogonia there is very clearly a longitudinal splitting of the chromatic loops. In the spermatocytes of normal pigeons there is no satisfactory evidence other than the remarkable decrease in the number of chromosomes to indicate the exact nature of the division but in hybrids, owing to certain exaggerations which frequently occur, especially in hybrids from very distinct species, some very significant facts come to light. As we shall see later, in many hybrid pigeons, the pairing necessary to the formation of bivalent chromosomes comes about with more or less difficulty or not at all. The chromosomes from the respective maternal and paternal species seemingly strive to maintain their own individuality in preference to fusing with a foreign plasma. The result is that they remain apart on separate spindles in some cases or unite feebly, or possibly sometimes fuse with members of their own species when two spindles are present instead of pairing with the chromosome from the opposite sex as they would do ordinarily. Where the two kinds of chromosomes refuse to unite, but divide in such a way that the paternal and maternal elements are set apart in separate cells, the effect is that of a qualitative reduction. In such exaggerated cases, then, what amounts to a reduction occurs in the primary spermatocytes, but this does not prove that the reduction ordinarily occurs here. If certain of the chromosomes can overcome their incompatabilities sufficiently to form bivalent chromosomes, as is true in the offspring from closely related forms, then their mode of procedure could be that of the ordinary bivalent chromosomes. To explain reversion and variation in the offspring of fertile hybrids, we are forced to the conclusion that a segregation of the paternal and maternal plasmas occurs in the germ cells, and the only chance for such a segregation, if the cells are of the normal type, is through the reduction division. It will be recalled that eight chromosomes were given to each secondary spermatocyte through the division of the primary spermatocyte, but only four appear when the cell is again ready for division; that is, the chromosomes have again united in pairs. In maturation the first division is ordinarily the equation division and the second the reduction division. If this be true in pigeons, then each of the eight chromosomes received by the secondary spermatocyte is bivalent. When these bivalent chromosomes fuse to form the four chromosomes of the secondary spermatocyte, then the latter must each be quadrivalent. A transverse division of such chromosomes could manifestly result in more varied distribution of the univalent components than the severing of an ordinary bivalent chromosome. The result would be greater diversity among the ultimate germ cells than would otherwise occur, and this would appear later in the form of greater variation among the offspring. The quadrivalent chromosomes could divide transversely in such a direction as to separate maternal and paternal chromosomes. This would seem to be the prevalent mode of division in hybrids, provided their respective parental qualities have not already been set apart in the primary spermatocytes. If the division of the quadrivalent chromosome be a transverse division at right angles to the direction just indicated, each ultimate germ cell would include both maternal and paternal chromatin. Both kinds of division occur, perhaps, in normal forms - possibly both kinds in the same cell. It can be seen at a glance that the number of possible combinations of qualities from egg cell and sperm cell at the time of fertilization would be considerably increased if both kinds of division take place. Offspring could possess the qualities of one grandparent on the paternal side to the exclusion of the other or exhibit the characters of each in equal proportion or in varying proportions. The same would be true, of course, with respect to the grandparents on the maternal side, provided the maturation of the egg has brought about a distribution of chromosomes similar to that seen in the spermatozoon. The fact that two lines of ancestry are merged in ordinary breeding complicates the matter, but it can readily be seen that such an arrangement of the chromosomes for the last division as is here indicated would add to the chances for variation in the offspring. The possibility presents itself then, that the second fusion of chromosomes as seen in the secondary spermatocytes is for the purpose of producing greater variation. This interpretation is, of course, as yet little more than a suggestion, and it is offered merely as an interesting possibility and as the only explanation the writer has been able to conceive of concerning the remarkable diminution in the number of the chromosomes from sixteen to four. |
TRANSFORMATION OF THE SPERMATID INTO THE ADULT SPERMATOZOON.
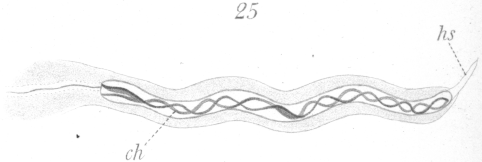
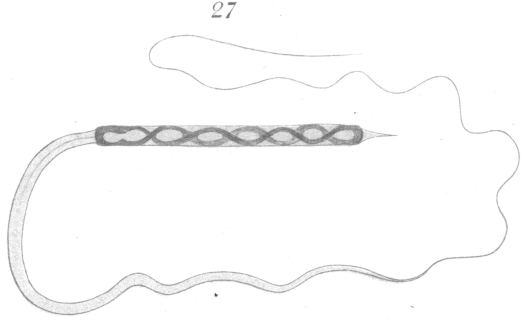
An adult spermatozoon as it exists in the vas deferens is shown in Fig. 27. The head is long and narrow and is intensely stained by nuclear dyes. Favorable preparations show faintly a spiral or vesicular arrangement of part of the chromatin in the interior. At the anterior end of the head is a slender finepointed head-spine. The head at its posterior end connects directly with the long cytoplasmic tail. No middle piece is visible. The tail and head-spine are very difficult to observe accurately, and but little of the details of their structure could be worked out. The only satisfactory way to gain a knowledge of the spermatozoon at all, in fact, is through a study of its development. Ballowitz {1. Ballowitz, E: Untersuchungen ueher die Struktur der Spermatozoon Arch. f. mikr. Anat., XXXII., 1888.} in his description of the spermatozoa of birds is somewhat in error in regard to the spermatozoa of pigeons (cf. his drawing Fig. 91,, Plate XVII.). He has seemingly mistaken the head for the tail.
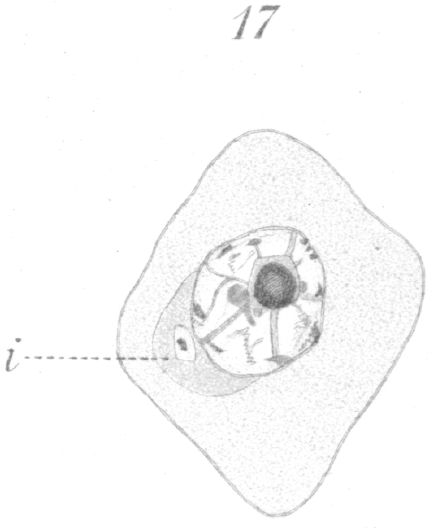
Before the spermatid begins its transformation, it is quiescent for a considerable period of time if one may judge by the large number that is nearly always observable in any section. When once the change begins, it is probably completed in a relatively short time. The spermatid, in the resting condition after the last division, has a very characteristic appearance (Fig.17). The nucleus is round and bears most of the chromatin at its center in the form of a large sphere which from optical section seems to be hollow. Fibers of linin radiate from the central mass to the nuclear membrane which is lined by a thin shell of chromatin. Not infrequently several small spheres of chromatin may persist at the center of the nucleus instead of one large one. The centrosome lies to one side of the nucleus imbedded usually in the sphere. The cell membrane is at times very indistinct.
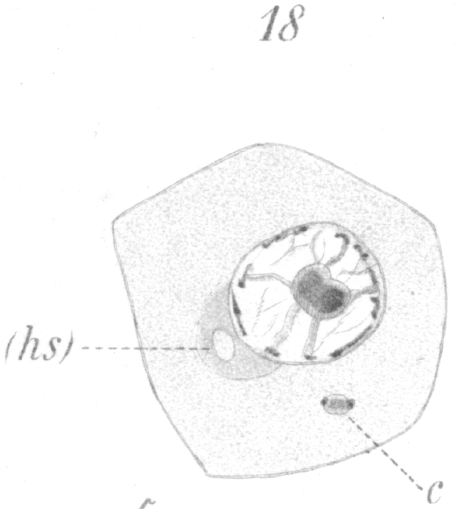
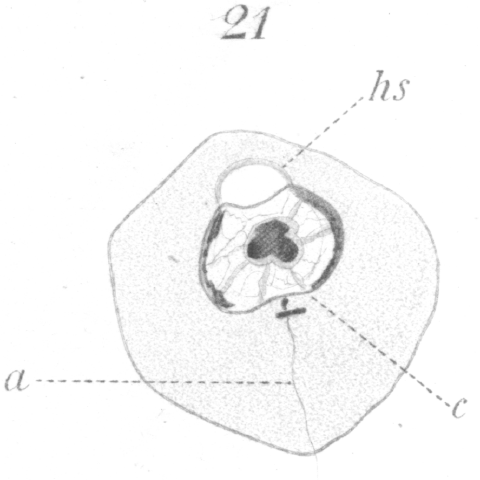
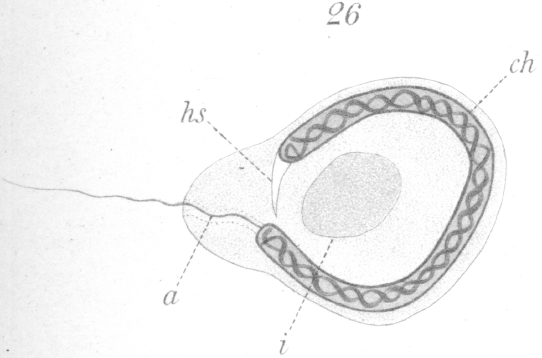
The first change to be observed is in the centrosome. It divides and moves out of the sphere and further away from the nucleus. A vacuole-like area persists in the sphere, marking the spot where the centrosome lay (Fig. 18, hs.). The two new centrosomes move apart for a short distance, but remain connected by a barrel-shaped sheath or mass of material (Fig. 18, c.). In the meantime the nucleus gradually approaches that part of the cell wall furthest removed from the centrosomes.
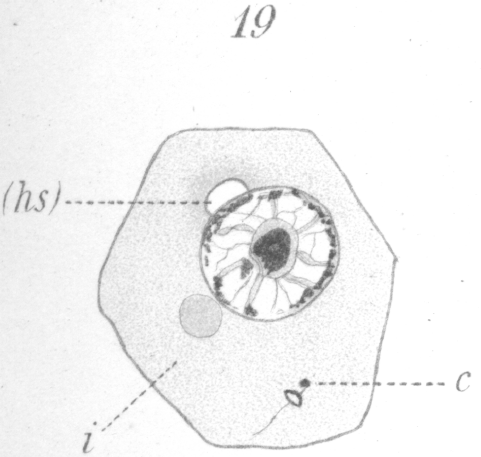
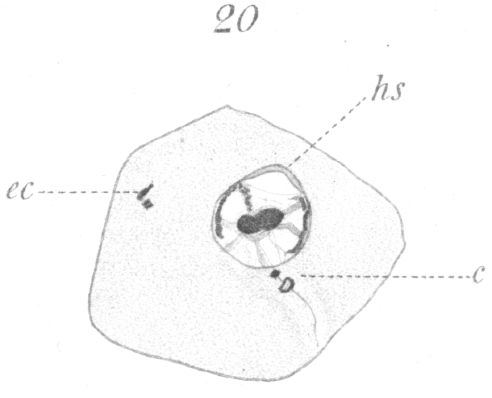
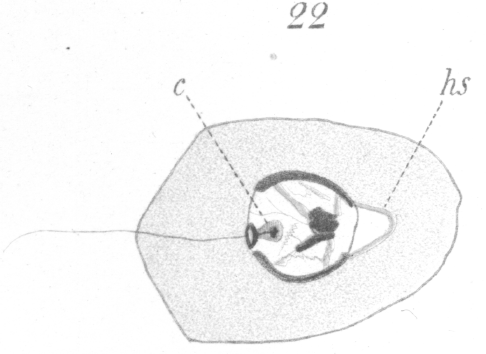
The next noticeable change is the disappearance of the barrel-shaped connective and the existence in its place of a very delicate fibril uniting the two centrosomes. One centrosome has enlarged, moreover, and if favorably situated for observation, can be seen to be a complete ring (Fig. 19). The connecting fiber just mentioned grows rapidly and is soon visible as a thread passing from the smaller centrosome back through the ring which it touches at one side, and continuing finally outside the cell (Figs. 20, 21). The vacuole or bubble-like area of the sphere gradually moves around the nuclear periphery in the meantime until it comes to lie at the pole opposite the centrosomes (Figs. 19-21, hs.). The centrosomes next approach the nucleus and so orient themselves that the smaller one points toward it. As they draw nearer a slight invagination appears in the nuclear wall and the smaller centrosome moves into it (Fig. 21, c.). The smaller centrosome is seen finally to lie inside the nucleus. The ring becomes closely applied to the nuclear wall where the invagination occurred. The filament or axial fiber of the tail, as it will ultimately be, continues to elongate.
A close examination of this stage discloses the fact that the invading centrosome has seemingly carried with it into the
nucleus an enveloping mass of extra-nuclear material which, judging from its reaction to stains, is of the same nature as the
sphere substance. Taking this into account together with the fact that in the adult spermatozoon there is no visible middle
piece, it seems probable that this internal centrosome together with its surrounding mass of cytoplasm may perhaps later develop into what corresponds to the middle piece of other spermatozoa. But here, since the material lies within
the nucleus, it is obscured by the rearrangement and crowding back of the
chromatin mass when the long head begins to form. Nothing further could be determined concerning the centrosomes as they
become lost to view at this point.
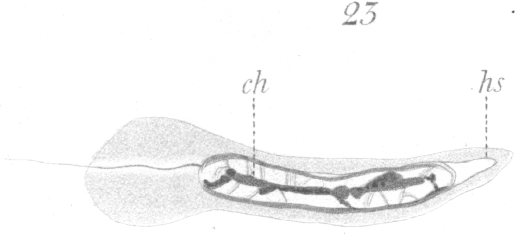
At this stage of development there is no trace of the sphere in the cytoplasm. The bubble-like structure mentioned above which originated in it, lies at what may now be designated as the anterior end of the developing spermatozoon. A flattening of the nuclear wall has occurred at the point of contact with the bubble and the latter becomes closely affixed or welded, as it were, to the nucleus (Figs. 20-24, hs.). The bubble-like portion develops ultimately into the head-spine (Figs. 22-27).
Soon after the centrosome has penetrated the nuclear
membrane, the elongation of the nucleus to form the head begins. At first only the anterior and posterior ends extend, but in a
short time the whole nucleus begins to narrow. Concomitantly, the chromatic mass at the center begins to sprout out both
anteriorly and posteriorly, and form a central core to the rapidly lengthening nucleus (Fig.
23, ch.).
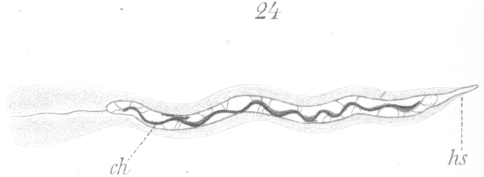
As the process of elongation continues in the nucleus, a
narrowing of the sides occurs to some extent, but when one takes into account the enormous elongation that takes place
together with the relatively slight diminution of the transverse diameter, it becomes evident that there must be considerable
increase in the volume of the nucleus.
|
The facts which appear to be of primary importance as set forth in the foregoing pages are briefly as follows: 1. The usual four types of cells exist in the genesis of the pigeon spermatozoon, viz :- spermatogonia, primary spermatocytes, secondary spermatocytes and spermatids. 2. Sertoli or nurse cells are present. 3. There is a curious ejection of part of the chromatin material from the nuclei of the spermatogonia just prior to their last division. 4. Sixteen loop-shaped chromosomes occur in the spermatogonia. They split longitudinally in division. 5. The last division of the spermatogonia results in the production of cells which, through a process of growth, become the primary spermatocytes. 6. Synapsis occurs in the primary spermatocytes through which a pseudo-reduction of the chromatin takes place. In the division of the spermatocyte only eight chromosomes appear, but they are in the form of heavy rings or vesicles and are evidently bivalent. 7. During the synaptic phase there is a marked drifting the of the chromatin to the side of the nucleus in contact with the sphere. Some of the contents of the nucleus apparently pass out into the sphere. 8. During division the eight chromosomes which are incased in capsules of linin break transversely, and as they move apart they remain connected by threads of the linin casing. These threads form the interzonal fibres. 9. Intermediate bodies at the equator of the interzonal fibres are present after all divisions. 10. The secondary spermatocytes result from the division of the primary spermatocytes. Some, at least, go into a resting stage, but it is of very short duration. 11. In the division of the secondary spermatocytes only four chromosomes appear. They are of the same size and shape as those of the preceeding division. It is suggested that they are quadrivalent. 12
.
If chromosomes are to be considered as differentiated
into qualitative areas, then a qualitative reduction apparently
occurs. It is possible that the chromosomes of the secondary spermatocyte are
quadrivalent. This would probably result in
a greater amount of variation in the offspring than would follow from the division of an ordinary bivalent chromosome. 17. In the transformation of the spermatid, the first perceptible change is in the centrosome. It divides and one of the resulting centrosomes enlarges and becomes ring-shaped. 18. The axial filament of the tail first appears as a thread connecting the two centrosomes and later continues backward through the ring-like centrosome and out of the cell. 19. The smaller centrosome together with material of cytoplasmic origin finally comes to lie inside the nuclear membrane and perhaps gives rise ultimately to a middle piece which becomes obscured by a covering of chromatin, and consequently appears to be absent in the adult spermatozoon. 20. The nucleus elongates to form the long head. It has a central core of chromatin in the form of a spiral filament which later splits to form a double spiral. 21. The head, during the later stages of development, undergoes a very great contraction but the spiral arrangement of the chromatin still persists in a modified form, constituting a series of eight vesicles apparently which may possibly be eight univalent chromosomes. 22. The head spine originates from a bubble-like mass of material which arises in the sphere. |
B. THE SPERMATOGENESIS OF HYBRID PIGEONS.
GENERAL CONSIDERATIONS.
It is a remarkable fact that no attempt has been made so far to investigate carefully the spermatogenesis or ovogenesis of hybrid forms. In all the mass of literature discussing or touching upon hybridism, so far as I have been able to ascertain, there has been in no instance an approach to a thorough study of the germ cells. Yet almost every writer states that through the study of hybrids, we have perhaps the best opportunity for gaining a clew to many of the most vital points in the great problem of heredity. A number of investigators have remarked that in certain instances the anthers, ovary, or testes as the case might be, were defective, and have let the matter go at that. There is practically no divergence from the normal type in the general method of procedure for the formation of spermatozoa in hybrids, hence it would be a needless repetition to go into all of the minutiae of development again. It will be taken for granted that the reader has determined the general plan of spermatogenesis from the first part of this paper. Only the modifications and irregularities which mark the hybrid will be considered here. The term hybrid is used in conformity with the definition given in the Century Dictionary which pronounces as hybrids such animals or plants as result from a cross of two forms noticeably different. Unsatisfactory as the definition is, it seems to be as accurate as it can be made. The objection, of course, is that it is uncertain how noticeably different two forms must be to be regarded as having individualities distinct enough to produce hybrid offspring when crossed. In the pigeon, some crosses are fertile, others are not. The sterile birds show a greater or less degeneration of the germinal cells. In the hybrid forms studied, the general rule seemed to be that the more divergent the parent forms, the more marked was the degeneration of the germinal cells.From parents which differ very widely in structure or habits, there is greater difficulty in securing female hybrids than male. The meaning of this is as yet a mystery. So far, I have been able to get but one female for microscopical examination. On the other hand, I have had six males, the offspring of very distinct species. These were all sterile forms. [Comment: Haldane's rule for hybrid sterility, enunciated 22 years later, noted that the heterogametic sex, which is the female in birds, is less evident among the offspring of crosses between diverging lines. See Haldane's Rule]. From the hybrid offspring of the common ring dove (Turtur risorius) and the white ring dove (Columba alba), a large number of sections were made for microscopical study. These two forms are perfectly fertile when crossed and the fertility of their offspring seems in no wise diminished. The latter are both fertile one with another, and with the parent species. The germ cells show some of the same phenomena as those of the the sterile birds, only in a much less marked degree. Offspring of the common ring dove when crossed with the white ring dove are brown in color. One member of the result pair is frequently a few shades lighter in color than the other. In the next or third generation there is generally a return to the original colors of the grandparents; one of the young is white, the other brown. Occasionally both of the young are brown or, less frequently, both white. There is a marked tendency for the white ones to be female and the brown ones male. Inasmuch as doves produce but two young at a brood, it would require a long period of time to secure numbers sufficient to arrive at satisfactory conclusions regarding the percentage of actual reversion to the original species. From the one character of color one might be led to infer that in the third generation there is in the majority of cases a reversion to the grandparent types, since the offspring are ususally one white and one brown. This conclusion does not necessarily follow, however, for we have no means of knowing whether the brown one gets its color from a return to the brown grandparent or directly from the parents of the second generation, both of which are brown. So far as the writer has carried his experiments, the indications are that on the whole there, are more brown than white birds in the third generation, and this points to the conclusion that in the brown birds we may have both intermediate forms like the hybrids of the second generation and forms which have reverted to the brown grandparent, as the white doves have seemingly returned to the white grandparent. [Comment: Guyer may have achieved the classical Mendelian ratios, with brown being dominant to white!] On the other hand, it does not necessarily follow that all white birds seen in the third generation have reverted entirely to the white ancestor. As has been mentioned, one of the doves of the original hybrid pair is often a lighter shade of brown than the other, hence the birds do not show the same degree of intermediacy. The birds of this generation, then, might mate in such a way that the offspring could exhibit the ancestral white while yet remaining intermediate in other characters. As we shall see in the conclusions from the study of the germ cells of hybrids, there are certain phenomena in the germ cells which apparently afford us a definite physical basis for the production of intermediate forms and for returns to pure ancestral species. From this basis there must necessarily be a greater number of intermediate forms in the offspring of hybrids than there are reversions to the respective ancestral species. Of the sterile hybrids, whether male or female, the sexual products were abnormal. As already stated, only one such female was examined. The facts observed concerning her will be briefly related later when the individual record of each form examined is given. Certain abnormalities exist in some phases of the fertile hybrid spermatogenesis as well as in the sterile forms, but they are much less pronounced and more restricted in range. The abnormalities of hybrid pigeons in general may be classified conveniently under three heads : (1) abnormalities in mitosis; (2) abnormalities in the structure of the spermatozoa; (3) degeneration of the germinal cells. Not all hybrids show these various irregularities in the same degree. All three classes of the phenomena just mentioned are observable in the sterile forms, but the fertile birds for the most part differ from the ordinary type only in the slightly irregular character of the mitosis. |
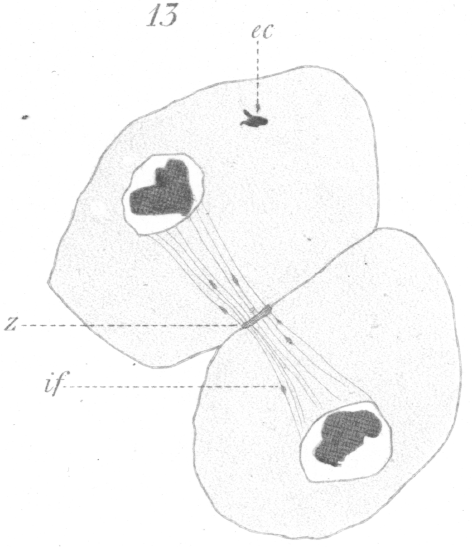
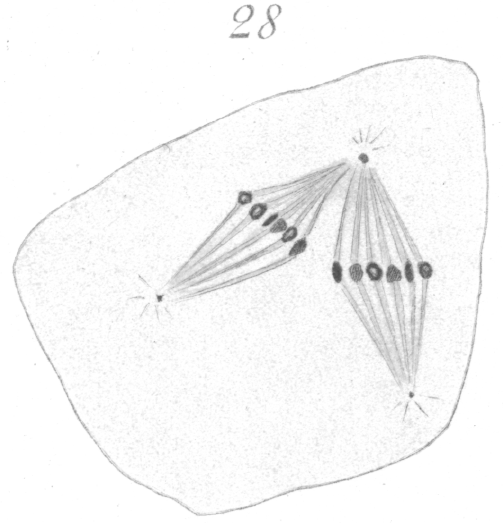
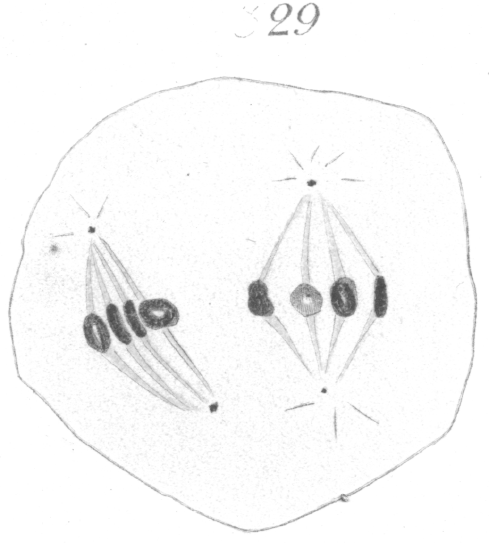
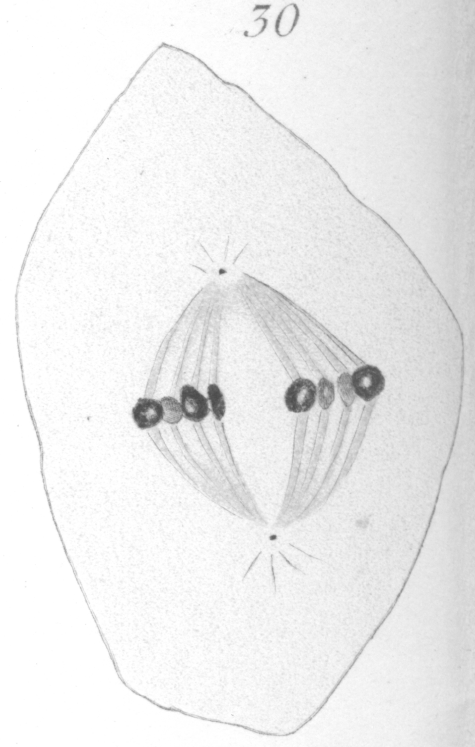
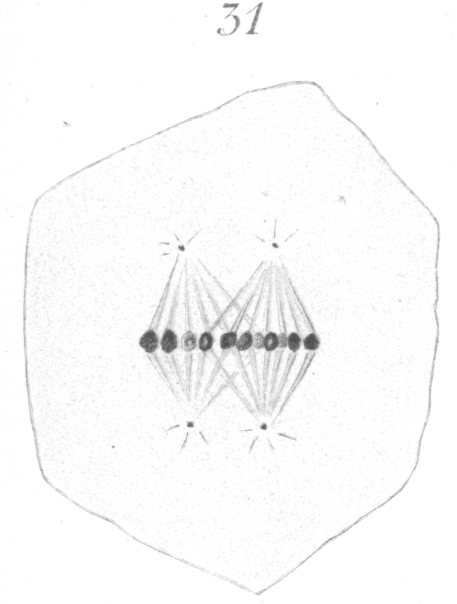
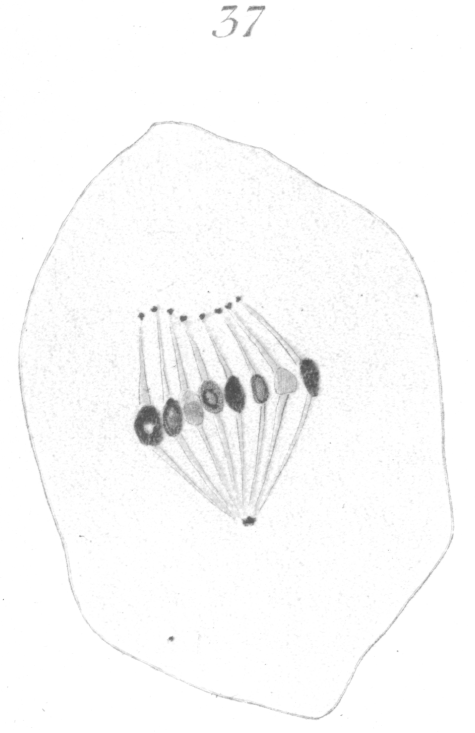
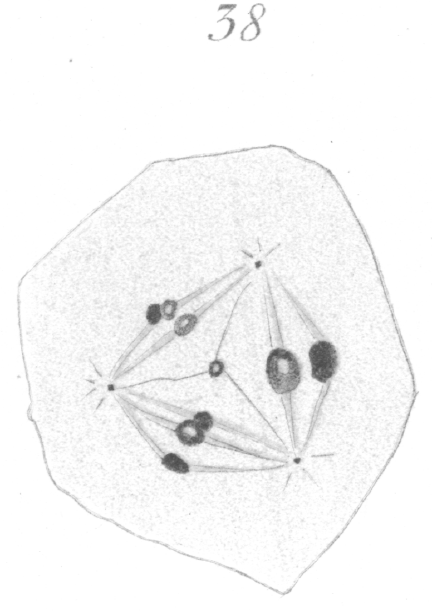
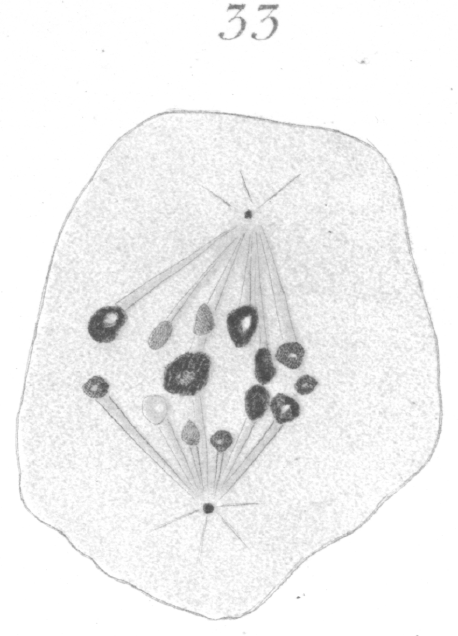
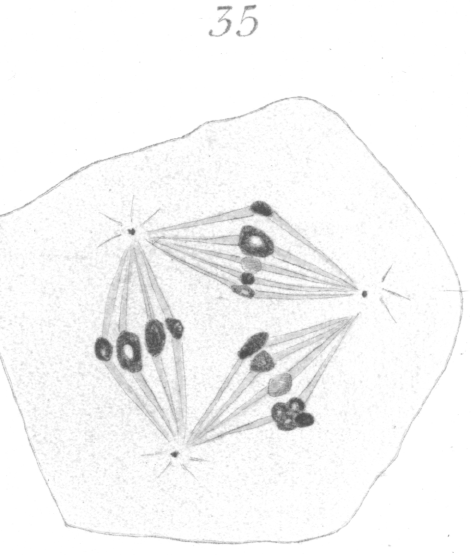
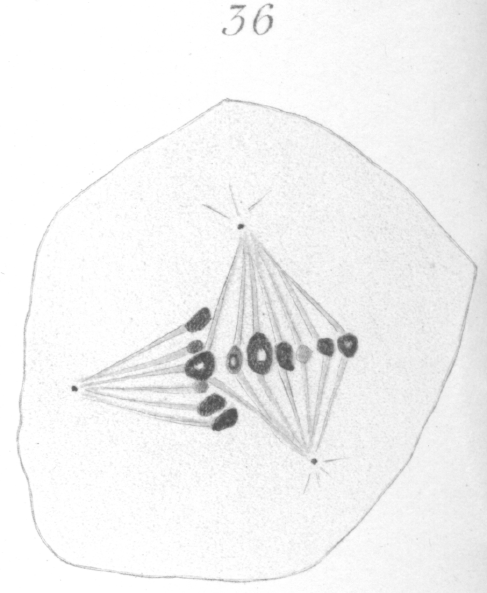
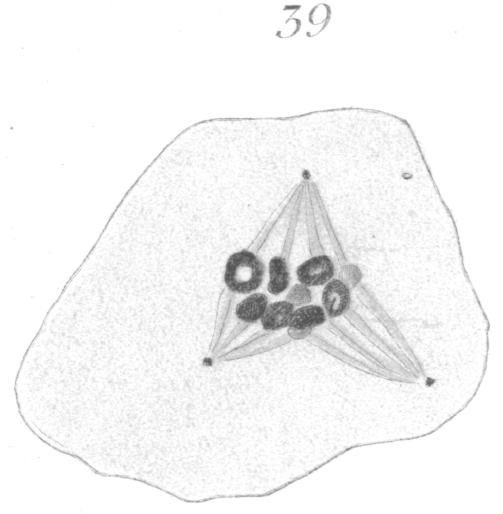
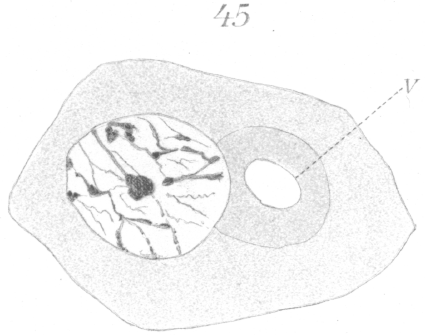
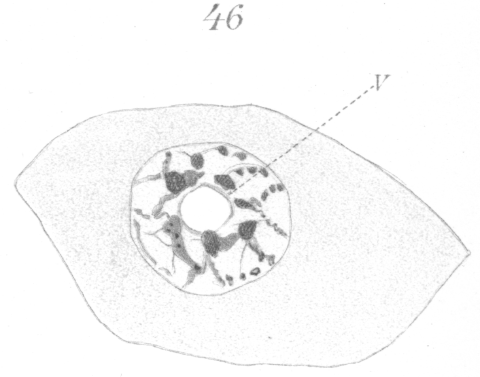
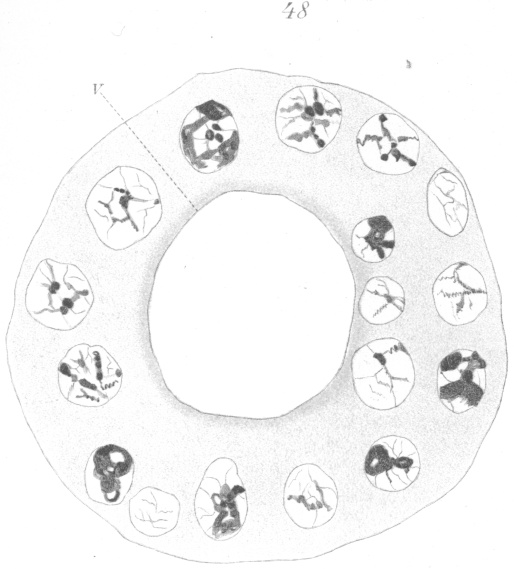
The abnormalities in mitosis are in the nature of multipolar spindles and asymmetrical division and distribution of the chromosomes (Figs. 28-39). These may exist independently one of another, or both may occur together in the same cell. They are more pronounced in sterile birds but may at times be seen in the fertile forms. In very many of the divisions of the primary spermatocytes one or the other, or both of these phenomena are seen. It is a curious fact that the multipolar spindles seem to be confined largely to the primary spermatocytes, and one is prompted immediately to associate the fact with the pseudo-reduction or formation of bivalent chromosomes which occurs normally at this stage of spermatogenesis. The irregularities in chromatin distribution are also seen for the most part in the primary spermatocytes.
Figs. 28-39 show some of the diverse forms of multipolar spindles that may occur. The tripolar types are by far the more common. Fig. 28 represents perhaps the most prevalent structure. It was not unusual to observe two spindles in one cell as shown in Fig. 29.
It is very easy to see how two independent spindles could be derived through a slight modification of such types as Fig. 28, 30 or 31. When two such spindles exist independently in one cell, they may each bear a small number of the large bivalent ring form chromosomes as in Fig. 29, or a greater number of small apparently univalent chromosomes.
In the first instance, judging from the general appearance of the spindles and the small number of chromosomes, it is
possible that the phenomena may indicate a precocious division
of the secondary spermatocytes; that is, it seems as if the divisions of both the primary and secondary spermatocytes have
been crowded into one period.
In the second case, which is the more usual, it appears rather that a synapsis or
pseudo-reduction has not occurred and that the chromosomes have retained
their univalent character and remained in two groups, each with a separate spindle.
More rarely both large and small
chromosomes appear on one or both of these spindles.
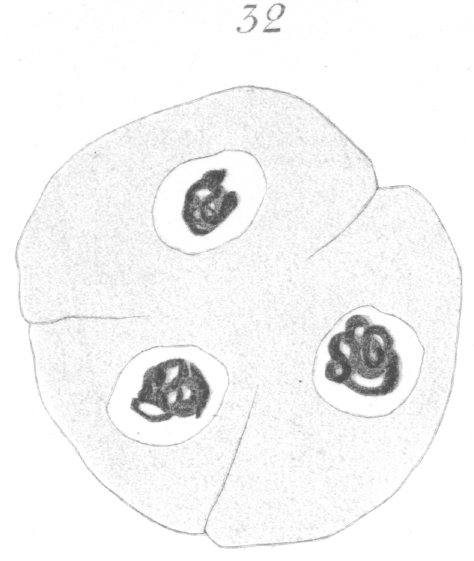
What has just been said regarding the chromatin arrangement where two separate spindles occur is equally applicable to the multipolar forms, only there is generally more variation in the size of the chromosomes. In a tripolar type like Fig. 28, the chromosomes are commonly of small size and are numerous. Fig. 32 shows the result of a division by means of a tripolar spindle; the three nuclei are passing into a resting condition.
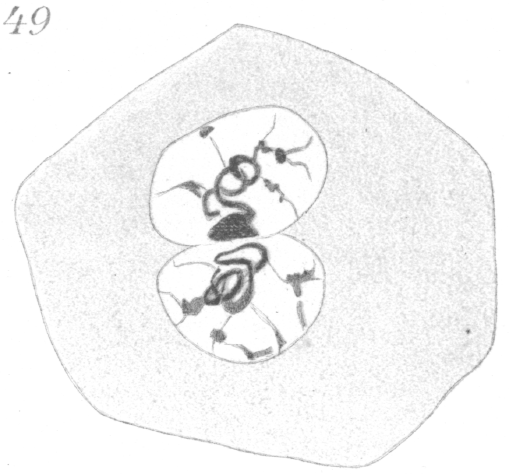
Fig. 31 shows a quadripolar type that is occasionally found. In a very few instances two nuclei were present in the primary
spermatocyte and it seems probable that such cells in division give rise to the multipolar spindles bearing an excessive amount
of chromatin which are sometimes seen.
Fig. 37 represents a case in which each fiber of the unusually
loose spindle seems to terminate at one end in a small centro
some-like dot or granule. Fig. 38 shows a tripolar spindle, possibly of a secondary spermatocyte, where there is much variation in the size of the chromosomes.
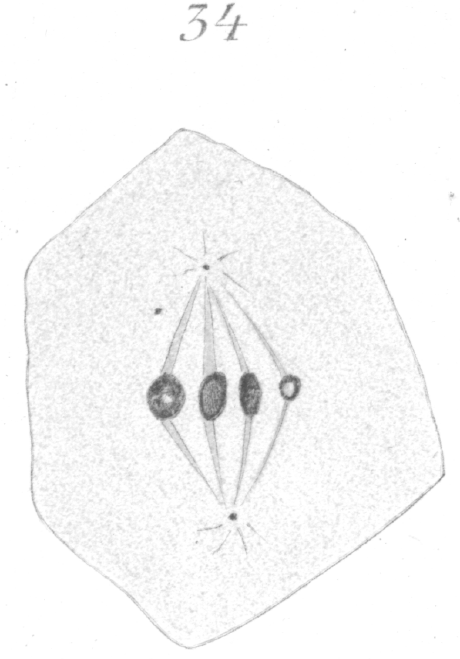
Then again, and more especially in the secondary spermatocytes, one of the chromosomes may be very minute (Fig. 34) as if part of its material had been lost in the preceding division.
In two or three instances in the division of the primary
spermatocytes, one of the large chromosomes was observed to be made up of four small rings
or vesicles (Fig. 35). It is possible that this indicates a demarcation into tetrads, visible in hybrid material because the firmer fusion
which occurs normally at this stage is lacking.
These irregular distributions of chromatin seem to affect
the size of the spermatozoa in the hybrid. There is apparently
a greater variation in the size of the spermatozoa than in normal pigeons, although the evidence is not as conclusive as is desirable, because killing reagents do not affect all
spermatozoa similarly.
However, where the same methods have been used on the spermatozoa of hybrid and of normal birds, there is certainly much more variation in length among the former. It was not uncommon to find giant forms three or four times the size of the ordinary ones, as well as unusually small ones. This extreme variation in the size of the spermatozoa was seen, however, only in hybrids from very different species, and such birds are, for the most part, sterile.
The objection arises, of course, that it is questionable if
irregular mitoses result in the formation of spermatozoa. When we take into account, however, the great frequency with which
these irregularities occur in some hybrids, and the fact that enormous numbers of spermatozoa may, nevertheless, be formed, it seems certain that many of them do mature into spermatozoa.
The irregularity in the formation of the spindle, moreover, does not indicate necessarily that the succeeding cells will be abnormal. It simply means, as we shall see, that
in all probability there is an incompatibility between the chromosomes from the two different species which, in some cases, prevents them from uniting for the pseudo-reduction that normally
occurs at this time. In
the other primary spermatocytes the same tendency must be present, and even if pairing of the chromosomes does occur, the succeeding division doubtless severs
the two plasma again and sets them apart in different cells, just as would be the case in many of the cells which, with their double spindles, visibly manifest the antagonism. From
the division of a tripolar spindle three cells will result, of which the one containing the end of both spindles will probably
receive chromatin from both equators and consequently from
both parent species. Each of the other cells will contain exclusively chromatin from different ones of the two spindles,
hence, each will represent but one of the two parent species.
Where two complete and separate spindles occur, of course, it is possible that none of the four resulting cells will be of the
mixed type; each may be pure. The unequal division of
individual chromosomes
which sometimes occurs would tend to
introduce chromatin of the opposite species into many otherwise pure cells.
|
ABNORMALITIES IN THE STRUCTURE OF SPERMAT0ZOA.
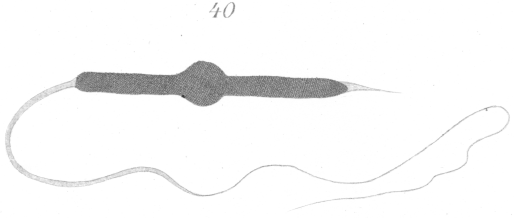
The misshapen spermatozoa that come under this heading
were present only in the sterile hybrids. In such forms there is
a curious varicosity or swelling about
the middle of the spermatozoan head (Fig. 40) that attracts the attention immediately
when the objects are examined under the microscope. This enlargement seems to be almost universal among the
spermatozoa of sterile hybrids and is sufficient of itself to produce sterility, for such a malformation would prevent its possessor
from entering the egg. In a very few instances what appears
to be a normal spermatozoon can be observed among the deformed ones and it is possible that if these reached a suitable
egg, fertilization might result. Although the odds against them
are very great, there is no reason apparent why they should not occasionally reach an egg and fertilize it.
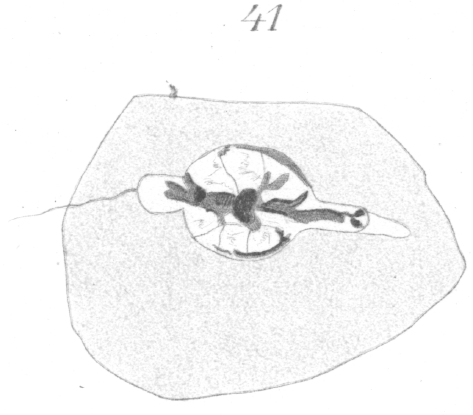
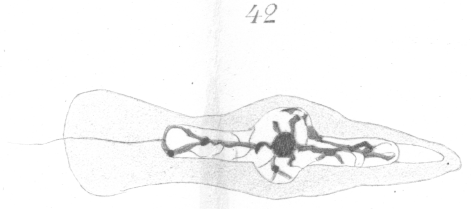
A study of the development of the spermatozoa reveals the fact that the bead-like enlargement results from the incomplete development of the nucleus to form the head. The two ends of the nucleus sprout out like the filament of a germinating spore and grow for a short distance, but the remainder of the nuclear wall retains its original form and position (Figs. 41, 42). The arrangement of the chromatin is very irregular. A deeply staining mass is visible in the bulb-like swelling, from which thick threads spring out forward and to the rear (Figs 41, 42).
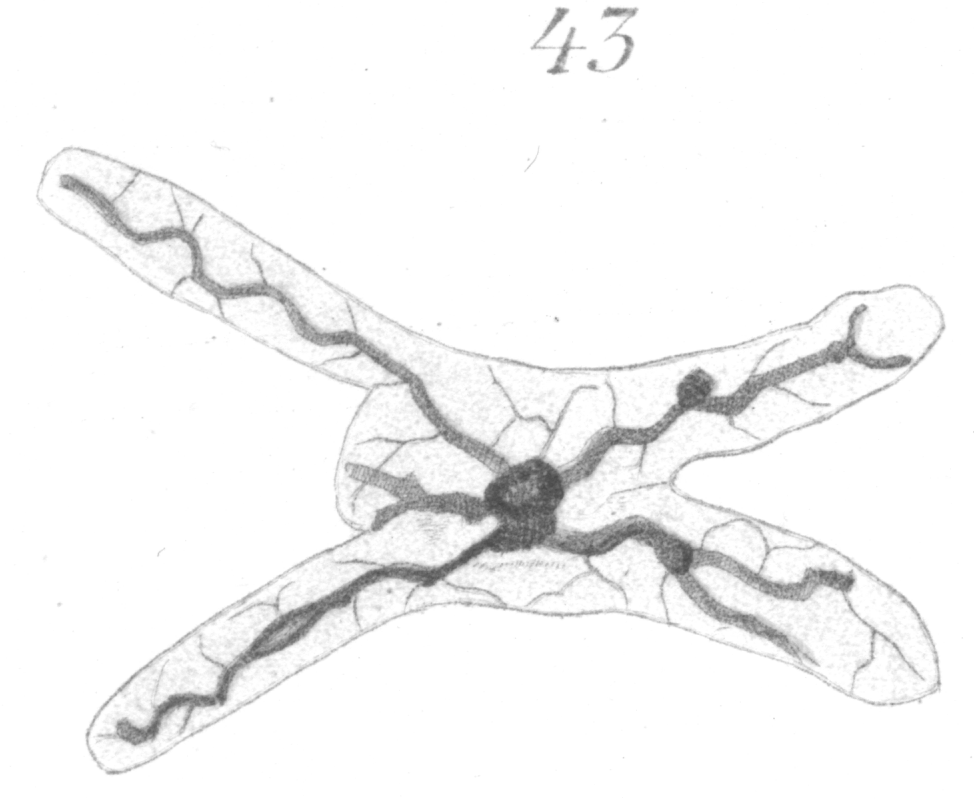
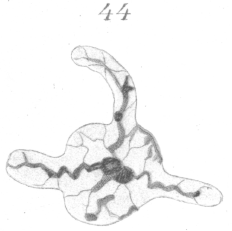
Figs. 43 and 44 show irregular forms of heads in which the protrusion or outgrowth of the nuclear wall has been in other directions than in the longitudinal axis. Such extreme forms are comparatively rare.
The deformed types, in which development is nearly complete, pass through practically the same stages as do the normal spermatozoa in their transformation from the spermatids. They may come to posses both head-spine and tail (Fig. 40), though it very frequently the tail is incomplete. In rare instances more than one tail appears in process of formation in the same cell, although no completely developed forms with two tails were observed. |
||||||||||
DEGENERATION OF THE GERMINAL CELLS.
Degenerative processes were in progress in the testes of all the sterile forms, but were most pronounced in hybrids between very divergent species, or between unlike hybrids from birds which were themselves descendants of fertile hybrids. There were some such extreme cases of degeneration that only the layer of cells lying along the wall remained in the tubule. Where such a degree of degeneration exists there is, of course, no approach to the formation of spermatozoa. There is often a strong invasion of wandering cells into the tubules, especially where the degenerative activity has become extensive. The interspaces between such tubules are also usually packed with cells which have much the same appearance as white blood corpuscles. Little clumps and globules of deeply staining cytoplasm are scattered about among the cells within the tubules. The oval nuclei of the Sertoli cells are also generally to be seen in varying numbers. In some places it really looks as if the germinal cells themselves lose their cell walls and characteristic appearance and become leucocytes, but this point will require very careful study before any definite conclusions can be reached. In other tubules the original cells, or cells which have wandered in, settle down and take on exactly the appearance of the large stroma cells which are ordinarily present outside the tubules. The cytoplasm of such cells has a peculiar alveolar like appearance that is very characteristic.
In birds where the degenerative processes are not so excessive, the tubules may yet contain large numbers of the germinal cells, many of which are in various stages of decline.
The primary spermatocytes seem to be the most susceptible to
decay. Fig.
45
represents a common example where the nuclear contents have a watery and disintegrated appearance and
the sphere substance is marked by the presence of a large vacuole in its center.
Fig. 46 represents another common type
in which one or more vacuoles appear in the nucleus. In Fig. 47 the spermatocyte has evidently prepared for division, as
evidenced by the eight chromosomes, but for some reason was unable to proceed. The chromosomes have remained intact
but have scattered throughout the cell.
Fig. 48 represents a type of very frequent occurrence. A number of cells have run together to form a giant cell in the center of which is an enormous vacuole. This last type is one very similar in appearance to the giant cells found in many pathological tissues.
Fig.
50
shows a degenerating cell in
which there are four groups of chromosomes. Each group is made up of four
chromosomes. This curious arrangement is
due probably to the fact that a cell like Fig. 29 has divided, but has ceased its activity
immediately after division before the
new nuclei had time to develop.
|
THE INDIVIDUAL HISTORIES OF THE STERILE FORMS
|
Before leaving the subject of the abnormalities in the hybrid forms, it seems desirable to run over briefly the individual cases that have been studied. The most important facts only are touched upon. The birds are described in no particular order. I. Age, 14 months. Parentage: male, tumbler; female, brown ring dove. The testes were large and from an external point of view fully developed. The ducts and genital papillae were normal. Degenerative processes were going on in many of the tubules but not so extensively as to prevent the formation of innumerable spermatozoa of the curious beaded type represented in Fig. 40. Giant cells (Fig. 48) were numerous as were also many vacuolated spermatocytes of the type represented in Figs. 45 and 46. Multipolar spindles and asymmetrical distribution of chromatin were very frequent in division. II. Age, 18 months. Parentage: Male, black tumbler; female, brown ring dove. Much the same condition prevailed as in Hybrid I. Not so many spermatozoa were formed and the degeneration was more marked. This might be due to the fact that the bird was older than the first. III. Age, 13 months. Parentage: male, a hybrid from a type white and brown ring dove; female, homer. The testes were very small. The vas deferens and genital papillae were normal. In the testes degeneration was extreme. Many of the tubules had lost nearly all of their germ cells. Numerous leucocytes were present. In some of the less affected tubules an occasional spermatid had become partially transformed into a spermatozoon, though no adult spermatozoa were found. In some tubules stroma-like cells were present. IV. Age, 11 months. Parentage: male, wild passenger pigeon; female, brown ring dove. The testes were of medium size. Inspection showed about the same conditions as existed in Hybrid III, although hardly as extreme. V. Age, 23 months. Parentage: male, wild passenger pigeon; female, brown ring dove. This bird was thought to be a female up to the time it was killed. It was mated with two different males and always played the role of female. The testes were found to be extremely small. The vas deferens was poorly developed and tapered off to a fine thread at the cloaca. The genital papillae were rudimentary. The degree of degeneration which prevailed surpassed that found in any of the other males. In many tubules only the peripheral layer of spermatogonia persisted; some of these, however, still showed mitotic divisions. Other tubules had become almost completely replaced by connective tissue. Not all tubules were equally affected. In some several layers of cells yet remained, but in dividing nearly every cell exhibited the spermatogonial type of chromosome and spindle. In a very few cases the regular spermatocytic type of chromosome was to be seen. VI. In connection with the male hybrids, it is perhaps well to mention another peculiar pigeon which was obtained from a dealer and whose antecedents are consequently unknown. It may have been a crossed form from two different varieties of dove-cot pigeons. The color markings were unusual. The neck and head showed numerous irregular splashes of white arranged in no definite pattern. There were extensive abnormalities in the testes. The spermatozoa were of the varicose type, and the divisions of the spermatocytes were characterized by multipolar spindles. Whether or not the bird was a hybrid could not be determined, but the case is an interesting one from whatever standpoint it is considered. Were it not a hybrid form, then there are other causes which produce effects in the germ cells resembling in appearance those of hybridism. This suggests the possibility of producing artificially irregularities in the division of the germinal cells. If such could be accomplished, it would be of great importance in determining whether or not variability can be induced in the offspring by means of chemical stimulus as well as through hybridization. VII. Age, 16 months. Parentage: male, common turtle dove; female, Japanese turtle dove. This was the sterile female already mentioned. The ovary was very rudimentary. The oviduct was of small diameter and but little convoluted. It opened normally into the cloaca. The ovary was no longer that of a very young bird, measuring only about 1 mm. in thickness and 4 mm. in length. It existed as a horseshoe shaped band which bent around the underlying mass of connective tissue, ganglia and blood vessels. Occasional small ova with rudimentary follicles of two or three cells were present. In only one instance was an egg found which showed much increase in size over the surrounding cells. It had attained to a diameter of 75 micra with a nucleus 37 micra in diameter. The ovum appeared much as a normal egg of the same size would except for the incomplete follicle. |
CONCLUSIONS FROM THE STUDY OF HYBRIDS.
|
It appears that in crosses of very divergent species the degenerative processes in the germ cells are at a maximum. In closely related forms like the brown and the white ring doves fertility is not diminished and the testes seem to be normal except for occasional irregularities in mitoses.The formation of multipolar spindles in division and the unequal distribution of chromosomes seen in many instances are among the most interesting phenomena presented. Such irregularities, however, occur to a slight extent in normal birds. A very careful study reveals the fact that they are more prevalent in the ordinary dove-cot pigeons than in pure breeds of doves which have bred true for many generations. That the irregularities of division in hybrids are due to degenerative processes going on in the tubules which give rise to deleterious chemical substances, is the first thought that presents itself. This seems a very acceptable idea from what we know of the effects of drugs upon cell division, but it does not account for the fact that the primary spermatocytes are the cells attacked in the great majority of cases. Asymmetrical mitoses, through the researches of Hansemann {1. Hansemann, D: Karyokinese end Cellularpathologie. - Berl. Kln. Wochenschrift. No. 42, 1891.}, Lustig and Galeotti {2. Lustig and Galeotti: Cytologische Studien ueber pathologische men- schliche Gewebe. - Beitr. Path. Anat. XIV, 1893.} and others, are known to be of very general occurrence in carcinoma cells and other pathological tissues, and Galeotti {3. Galeotti, G: Ueber experimentelle Erzengung von Unregelmassigkeiten des karyokinetischen Processes. - Beitr. z. patholog. Anat. u. z. Allg. Pathol., XIV, 2, Jena, Fischer, 1893.} and Schottlander {4. Schottlander, J: Ueber Kern and Zelltheilungsvorgange in dem Endotheil der enzandeten Hornhaut. - Arch. f. mikr. Anat. XXX, 1888.} among many others have determined that they may likewise be produced artificially in many tissues by treatment with dilute, solutions of various drugs, such as quinine, chloral hydrate, cocaine, potassic iodide and antipyrin. A hybrid offspring is really a compound of two very different individual plasmas, hence conflicting tendencies must necessarily have been induced within its body. Most of the abnormality in division is in all probability but an attempt of each plasma to assert its individual activity. But why does this effort become apparent only in the primary spermatocytes? The answer appears to be simply that there is no necessity for fusion of the components of the chromatin of cells except in the germ-cells at one stage of their maturation. Investigators have found that in maturation of germ-cells, a reduction of the ordinary number of chromosomes to one-half occurs. Before this actual reduction there is ordinarily a so-called pseudo-reduction in which the chromosomes fuse in pairs so that when the cell is ready for division, although only half of the regular number of chromosomes appears, each is really double (bivalent)and equivalent to two of the simple (univalent) type. In spermatogenesis this change generally comes about in the primary spermatocyte. Now, in hybrids it may be supposed that in the ordinary cells of the body, the chromosomes from the paternal and the maternal species lie side by side and carry on the customary functions of the cells but when it comes to an actual fusion of chromosomes to form the bivalent type necessary for reduction, the incompatibility of the two different plasmas renders the union incomplete or prevents it entirely. That the ordinary somatic cells of hybrids, either plants or animals, are under the influence of two distinct tendencies is well shown in the decided mosaic-like structures which frequently occur, the classic figure being to liken hybrids to warp and woof. To cite but one example, Macfarlane {1. Swingle, W. J. and Webber, H. J: Hybrids and Their Utilization in Plant Breeding. - Yearbook, Dept. of Agric., 1897.} found that a hybrid of the gooseberry and black currant instead of being a strictly intermediate type, really possessed side by side, organs characteristic of each parent; the leaves bore both the shield-shaped, oil-secreting hairs of the currant and the simple hairs of the gooseberry, though each hair was but half the size of the parent type. We may infer then that in Hybrid pigeons the univalent chromosomes from each of the parents may lie side by side in the ordinary cells of the body and divide normally, but when it comes to the period of fusion in the germ-cell, they will not unite to form the bivalent type or else they unite incompletely. The result is that in the primary spermatocyte instead of one spindle bearing eight bivalent chromosomes, a multipolar spindle, or not infrequently two separate spindles, bearing two groups of univalent chromosomes may appear. In cases where both large and small chromosomes are seen, it is necessary to suppose that a loose union has occured in some chromosomes. Whether the bivalent chromosomes formed under such conditions consist of chromatin from only one parent, or whether both parents are represented, there is no means of determining. The unequal divisions of the bivalent chromosomes of hybrids indicate that such chromosomes have in some way been rendered very unstable. In chromatin we have a substance which from all we know of its nature and actions, seems to be intimately bound up in the phenomena of inheritance. It is reasonable to suppose, therefore, that it constitutes at least a part of the material basis for variation in the germ. The question then arises as to whether there is any correlation between the distribution of chromatin as seen in the germ-cells of hybrids, and the marked variability which characterizes the offspring of fertile hybrids. If we accept the view that chromatin is a substance capable of varying in qualities in the different regions of the chromosome, then in fertile hybrids where irregular mitoses occur, the different germ-cells will certainly not be qualitatively similar after division and one would expect the offspring produced from such cells to be variable. That the chromatin of each parent species often retains its individuality, is indicated by the fact observed in many primary spermatocytes where two separate groups of the small or single type of chromosome exist. The division of such a cell into three or four, as the case may be, results in the formation of new cells, some of which will manifestly contain chromatin from only one of the original parent species and some, only from the other. Some of the spermatozoa then, will bear chromatin from only one of these species. In the offspring from such a cell one would expect a much closer return to whichever one of the parent forms it represented than in the offspring of a "mixed" spermatozoon. In discussing irregular divisions, however, it must not be forgotten that many apparently normal divisions of the primary spermatocytes also occur in all hybrids, and constitute by far the predominant kind of division in hybrids from closely related forms. Unequal distributions of chromatin cannot therefore play the most important part in variation or reversion. There seems to be no other interpretation, indeed, than that in the many normal mitoses of the bivalent chromosomes which occur, the chromatin of the father and of the mother is set apart so that the ultimate germ-cells are what might be termed "pure" cells; that is, a given egg or sperm-cell contains exclusively or at least predominantly qualities from one parent. The offspring from fertile hybrids of the same parentage might then be similar to the mixed type of the original hybrid, or revert to one of the grandparent types, dependent upon the chances of the various cells for union at fertilization. If a spermatozoon and an egg containing characteristics of the same species unite, then the reversion will be to that species; if a sperm-cell containing the characteristics of one species happens to unite with an ovum containing characteristics of the other species, then the offspring will be of the mixed type again. By the law of probability the latter will be the more prevalent occurrence, because there are four combinations possible, and two of the four would result in the production of mixed offspring, while only one combination could result in a return to one of the ancestral species. From the fact that in cases of apparently complete return to one parent type, characteristics of the other parent may nevertheless crop out from time to time in succeeding generations, it is evident that all of the germ-cells are not absolutely "pure." The occasional inequalities in the division of individual chromosomes, as already mentioned may account for this fact. Not infrequently in division more than half of one chromosome goes to one cell; thus, what might otherwise have been a pure cell will contain a few characters of the foreign species and these may appear later in offspring as variations of the intermediate type. Thus the actual number of intermediate forms will doubtless be augmented somewhat beyond the proportion indicated in the probability of combination where all the mature germ cells are pure. It is also possible that such mixed cells as result at one end of a tripolar spindle will introduce yet another factor for variation though tripolar spindles are too few in most fertile hybrids for this element to enter to any great extent. It is probable that the irregular distributions of chromatin, where such occur, have more to do with such succeeding offspring as show variation, and less with those which return to the specific types. In the latter case the chromatin of each species has frequently remained entirely distinct (in marked hybrids), or has normally separated again at the sundering of the bivalent chromosomes (in mild crosses) into the two original plasmas. Thus it is very obvious that most of the variation seen in the offspring of fertile hybrids is due to the union again of two "pure" germ-cells each of which represents a different one of the original parent species, but it is also an evident that inequality in the distribution of chromatin may enter as a factor. That irregular divisions cannot account entirely for reversions to grandparent types is very evident in the crosses of brown and white doves, where the irregularities are by far too few to equal the percentage of reversions. There is but one ultimate conclusion, then, namely, that the irregularity in division of the primary spermatocytes which appears in hybrids between very different species, is but an index to what occurs in ordinary crosses. In the latter, instead of separate spindles and non-fusion of chromosomes, a true union occurs, but the bivalent chromosomes ultimately divide in such a way that the respective plasmas occupy different cells. There is a separation of the paternal and the maternal chromosomes which had fused during synapsis. With regard to the question of the persistence of chromosomes, the evidence is becoming stronger every day that these elements do retain their individuality. In addition to the well known observation of Boveri {1. Boveri, Th: Ueber die Befruchtung der Eier von Ascaris megalocephala. - Sitzungsberichte d. Ges. Morph. n. Physiol. Munchen, II, 1887. Ueber partielle Befruchtung. Ibid., IV, 1888.} on the early stages of Ascaris, there are numerous other facts which point to the same conclusion. Ruckert {2. Ruckert, J.: Ueber das Selbstandigbleiben der vaterlichen and mutter lichen Kernsubstanz wahrend der ersten Entwicklung des befruchteten Cyclops-Eies. - Arch, f. mikr. Anat. XLV, 1895.} for instance, in his study upon the fertilization of cyclops, was able to follow the maternal and paternal chromosomes very distinctly in cleavage. Zur Strassen {3. Zur Strassen, O: Ueber die Riesenbildung bei Ascaris-Eiern. - Arch. Ent. wm. VII, 1898.} has shown that in the giant embryos of Ascaris which arise by a fusion of eggs, the number of chromosomes increases proportionately with the number of nuclei which unite. Again, to cite one more of the rapidly multiplying examples, perhaps the most brilliant confirmation is met with in the work of Herla {4. Herla, V.: Etude des variations de la mitose chez 1'Ascaride megalocephale. - Arch. f. Biol. XIII, 1893.} and of Zoja {5. Zoja, R.: Sullo independenza della cromatina paterna a materna nel nucleo dele cellule embrionali. Anat. Anz., XI, 1895.}. Both observers have shown that in the hybrid fertilization of Ascaris, if the egg of variety bivalens is fertilized with the spermatozoan of variety univalens, the three chromosomes thus brought together retain their individuality and reappear at each cleavage, at least to the twelve-cell stage. Zoja affirms that the paternal chromosome is of smaller size and is thus distinguishable from the two maternal chromosomes. The above interpretations are offered with the hope that they may perhaps lead to some clew concerning the real nature of the material basis of heredity. If the conception proves to be a true one, then it doubtless affords a key, among other problems, to the long-standing one as to why many plants will come true from slips or grafts, but not from seed. The reason may be sought in the pseudo-reduction period of the germ-cell. Plants such as the apple, for example, which do not come true from seed, are practically multi-hybrid. In the germ-cells there will be numerous incompatibilities due to the fact that the plant has been miscellaneously fertilized for a number of generations. In propagation by means of slips, the chromosomes lie side by side and divide in the ordinary way to construct and maintain the new body, so that it is practically a continuation of the old one; but when the time comes for maturation of the germ-cells, the lack of harmony between the various plasmas asserts itself, with the result that bivalent chromosomes are formed, which divide in such a manner as to segregate different sets of ancestral qualities. The resulting combinations in fertilization will give rise to seed many of which may possess dissimilar sets of qualities. Concerning the other abnormalities met with in the spermatogenesis of hybrids, about all that can be said is that the whole phenomena show lack of vigor in the development of the germ-cells, whatever this may mean. The deformed spermatozoa indicate want of sufficient vitality to push the development through to completion. The germ-cells start out apparently to perform their functions normally, but later succumb to the conflicting forces at work within their boundaries. In most instances where the species are very distinct, the hybrids can be obtained in the first place only, as it were, under protest [Comment: Hybrid inviability]; the breeding season must be at its height and everything in the most favorable condition possible. As to why the reproductive organs should be more susceptible to abnormal changes than other regions of the body, we appear have no clew. Darwin has pointed out repeatedly the curious parallel between crossing and the change produced by physical conditions. Animals and plants removed from their natural conditions are extremely liable to have their reproductive systems affected. Still he recognizes that sterility is incidental and not a necessary concomitant of hybridism. In some fertile hybrids, indeed, there seems to be an improvement on the parent form [Comment: Hybrid vigour.]. Thus Vernon {1. Vernon, H. M : The Relation between the Hybrid and Parent Forms of Echinoid Larvae. - Phil. Trans. Series B. CXC, 189.} found two species of sea urchins, which when crossed showed greater fertility and produced larger, stronger larvae than in direct fertilization. |
The above conception of the chromatin does not necessarily imply that it is of exclusive importance as the basis for hereditary transmission. There seems to be no sufficient reason for not regarding the cytoplasm likewise as an important factor. Connected with the possibility of the qualitative differentiation of the chromatin there arises another possibility, namely, that only the individual and the more variable characteristics are conveyed by the chromatin, while back of these in the cytoplasm is a general stock or plasma from which the more stable and constant form of an animal is derived. In other words, that there is a sort of double background of qualities to every organism which might perhaps be designated as general and individual. The cytoplasm provides the general substratum upon which all the later individual traits controlled by the chromatin are gradually built up. To illustrate in pigeons: what is recognized as the pigeon type or the general characteristics that mark a particular bird as a pigeon would constitute the first or general type [Comment: species specific characteristics], and those characteristics by which one recognizes different groups of pigeons or different individuals, would represent the second or individual type..[Comment: classical Mendelian characteristics] Such a conception, among other things, would simplify our interpretation of reversion to a distant ancestral form. With reference to the actual occurrence of such reversion it may be said that there are sufficient well authenticated cases on record to establish beyond a doubt its existence. Among others, Ewart {1. Ewart, J. C: The Penycuik Experiments., 1899.}, in a recent paper, gives some beautiful corroborative evidence.
To explain reversions, some investigators presuppose the existence of latent elements which have lain dormant for perhaps many generations and which in some unknown manner later reassert themselves. Others, on the contrary, maintain that the resemblance is produced through a recurrence of suitable external conditions, such as existed during the ancestral period. A third and apparently simpler possibility arises if we conceive of the cytoplasm as the more conservative, or stable, and the chromatin, the more individual and variable agent in the germ cell. Thus, if little chromatin were present, or if it were inactive for any reason, as, for instance, if certain qualities neutralized each other, then from the cytoplasmic portion would develop the general or ancestral type which is the same for all the kindred forms. The cumulative individual characters latest added to the chromatin would remain in abeyance and the result would be what is called reversion. No latent germs are necessary beyond the original fundamental ground substance of the type, for the reversion is not due to a reassertion of a latent germ, but to the suppression of the later more developed characters which are transmitted through the chromatin.
After having penned the above the writer was very much interested in noting that Ewart {2. Ewart, J C: The Penycuik Experiments., 1899, page xxviii.} voices much the same idea regarding reversion although he suggests no localization in the germ-plasm. Speaking of reversions in pigeons, he remarks:
A supposition that would be required to supplement the above conception is that the qualities of the nucleus after long continued recurrence must finally become stamped upon the cytoplasm, else there could be no evolution or development of the type itself. Our knowledge of nucleus and cytoplasm so far is, of course, too crude to give a justifiable basis for such an assumption beyond that of mere suggestion, but from what we do know of the interrelation of cytoplasm and nucleus, it would seem that perhaps this difficulty may not, in the end, prove insurmountable. According to such an assumption as the above regarding the specific nature of the chromatin, one would expect the early and more fundamental phases of development to be determined by the cytoplasm and only the later and more individual by the chromatin. The cytoplasm of the female, then, would probably give the general form, inasmuch as ordinarily little cytoplasm enters with the sperm. This would mean that a young hybrid would resemble the female but would come more and more toward the male type as it matured, after reaching a point where special individual characteristics begin to crop out. That is, the influence of the male would not be felt till the latter part of development. Let us now examine some existing facts in the light of this suggestion. Driesch and Morgan {1- Driesch, H. and Morgan, T. H: Zur Analysis der ersten Entwicklungsstudien des Ctenophoreneies. - Arch. Entwm. II, '95.} demonstrated that if part of the cytoplasm of an unsegmented ctenophore egg is removed, a defective larva will result; that part represented by the removed portion is incomplete. This result shows that the cytoplasm as well as the nucleus must be reckoned with in the problems of heredity. Again Driesch {2. Driesch, H.: Ueber rein-mutterliche Charaktere an Bastardlarven von Echiniden. - Arch. Entwm. VII, 1898.} records that the egg cell determines the rate of cleavage in certain Echinoderms when slow and rapid cleaving species are crossed. The mesenchyme cells, which are peculiar in each species, follow the egg cell parent type in the hybrid. Standfuss {3. Standfuss: Handbuch der palarktischen Gross-schmetterlinge. 2nd. Edition, Jena, 1896.} through experiments upon butterflies, shows that young hybrids at first resemble the female species, but with each succeeding metamorphosis diverge more and more toward the male species, until the hybrid comes to its final degree of intermediacy. The fact that denucleated fragments of eggs which have been fertilized by the sperm of another species give rise to offspring which resemble the paternal species does not indicate that necessarily that the conception under discussion is untenable. In For example, Boveri {1. Boveri, T'h: Ueber die Befruchtun.gs-and Entwicklungs-lohigkeit kernlo,ser Seeigel-Eier.-Arch. Entwm. II, 1895.} in fertilizing a denucleated egg of one species of sea urchin with the nucleus of another species notes that the resulting animal when completely developed, resembles the latter species. This is just what one would under the circumstances anticipate. We should expect the cytoplasm of of the egg to determine none but the first general stages. In as much as there is chromatin present from one form only, the paternal, the individual finally produced would necessarily resemble that form exclusively. The above statements are offered only in the form of suggestions. For a hypothesis to bear any probability of truth it must harmonize with existing facts, and there is none who will deny that existing facts in regard to heredity are far too few to admit of a dogmatic statement of any theory. Most suggestions are valuable, at present, only as they stimulate to the acquisition of more facts. |
|
The points of special interest in the spermatogenesis of hybrid pigeons are as follows: 1 . The general plan of spermatogenesis is not essentially different from that of normal pigeons.2. From forms not too distantly related, fertile hybrids are easily obtained. 3 Infertile hybrids, resulting from the crossing of very distinct species, are more difficult to secure. Females are rare. 4. All hybrid pigeons exhibit multipolar spindles and asymmetrical distributions of the chromatin in cell division. These irregularities are much more infrequent in fertile hybrids. 5. Sterile hybrids show in addition, a deformed spermatozoon and often a marked degeneration of the germinal cells. 6. The irregularities of division are confined for the most part to the primary spermatocytes. Likewise it is in these cells that the formation of bivalent chromosomes occurs normally. In hybrids, it would seem that the conflicting tendencies of the of one two parental plasmas frequently render the union of the single chromosomes to form the double (bivalent) types impossible or abnormal. There seems to be an attempt on the part of each plasma to assert its individuality. This visible incompatibility of the chromosomes from widely different species serves as an index to a kindred lack of harmony between the plasmas of more nearly related forms, so that even though pseudo-reduction does occur and normal division of the bivalent chromosomes follows, the identity of the individual species is still retained through the segregation of the maternal and paternal chromosomes into separate cells, which may be considered "pure" germ-cells (containing qualities of only one species). 7. Union of two cells containing characteristics of the same species would occasion a reversion to that species. Union of two cells representing each of the two original species would yield an offspring of the mixed type. The latter would predominate because of the greater probability of such union. Besides, through the mixing just indicated, variability may be due also in some cases to the not infrequent inequalities in the division of individual chromosomes, through which varying proportions of the chromatin of each species may appear in certain of the mature germ-cells. 8. Irregular divisions cannot of themselves account entirely for reversion and variations, because double spindles and irregularities in the formation of bivalent chromosomes are far too few to equal the percentage of reversions seen in such mild crosses as the brown and the white ring dove. One is forced to the conclusion expressed above, that the double-spindled and multipolar types of cells which occur in hybrids and between very divergent forms are but exaggerated images of a tendency which exists in the primary spermatocytes of normal appearance, which are to be found in all hybrids. 9. The above conception may likewise afford a clew to the problem of why certain plants will come true from slips or grafts, but not from seed. The explanation may be sought in the pseudo-reduction period of the germ-cell. 10. The deformed spermatozoa are due to the incomplete development of the nucleus of the spermatid to form the head. 11. It is suggested that the physical basis of heredity may be dual in nature, involving both cytoplasm and chromatin. HULL ZOOLOGICAL LABORATORY, The University of Chicago, May 10, 1900 |
BIBLIOGRAPHY.
|
BALLOWITZ, E
: Untersuchungen ueber die Struktur der Spermatozoan. - Arch. f. mikr. Anat.,
XXXII,
1888. HERLA, V: Etude des variations de la mitose chez 1'ascaride megalocephale. - Arch. Biol. XIII, 1893. JUEL, H. O: Die Kerntheilungen in den Pollenmutterzellen. - Jahrb. wiss. Bot., XXX, 1897. LENHOSSEK, M. v: Untersuchung ueber Spermatogenese. - Arch. f. mikr. Anat., LI, 1898. LUSTIG, and GALEOTTI, G: Cytologische Studien u.eber pathologische menschliche Gewebe.--Beitr. Path. Anat., XIV, 1898. MCCLUNG,
E. C: A Peculiar Nuclear Element in the Male Reproductive Cells of Insects. Zool. Bull. II, 4,
1899.
MOORE,
J. E. S: On the Structural Changes in the Reproductive Cells during the Spermatogenesis of Elasmobranchs.
- Quart. Journ. XXXVIII,
1893. WILCOX, E. V : Spermatogenesis of Calloptemus femur-rubrum and Cicada tibicen. - Bull. of the Mus. of Comp. Zool., Harvard Coll., XXVII, i, 1895. ZOJA, R: Sullo independenza della cromatina paterna a materna nel nucleo delle cellule embrioli. - Anat. Anz. XI, 1895. ZUR STRASSEN, O: Ueber die Riesenbildung bei Ascaris-Eiern. - Arch. Entwm., VII, 1898. |
End of Thesis. This copying was made possible by a generous loan to DRF of an original copy by the Library of Brown University.
![]()
For Bateson and Saunders (1902) "Two Levels of Genetic Information" (Click Here)
Next: For Winge (1917) on Chromosomes, Polyploidy and the Origin of Sex (Click Here)
Return to Main Guyer page (Click Here)
Return to Bungener & Buscaglias' analysis of Guyer (Click Here)
Return to Evolution Index (Click Here)
Return to HomePage (Click Here)
![]()
This page was created in April 2004, and was last edited on 08 Sep 2011 by D. R. Forsdyke