Michael F. Guyer's Contribution
Patrick Bungener & Marino Buscaglia
Institut d'Histoire de la Medecine et la Sante et Departement d'Histoire
et Philosophie des Sciences, UniversitE de Geneve
Hist. Phil. Life Sci., 25 (2003), 27-50
[Reproduced here, without comment, by DRF with the permission of the authors. The publishers in 2003 were Taylor & Francis Ltd. On being contacted for permission, they referred DRF to the current publishers, Giannini Editore of Naples. But the Managing Editor of HPLS, Christiane Groeben , informed DRF (Sept 1, 2011) that copyright authority was transferred to the Statione Zoologica Anton Dohrn, and did not "see any reason why you should not be allowed to put the paper on the website, once you have the permission from the authors". ]
Introduction
Historical_background_of_the_Sutton-Boveri_chromosome_theory
Guyer's_dissertation
Guyer's_hypothesis_and_the_rediscovery_of_Mendels_principles
Reception_of_Guyer's_hypothesis_by_the_contemporaries
Conclusion
ABSTRACT - This paper examines the contribution of the PhD dissertation of the American cytologist Michael F. Guyer (1874-1959) to the early establishment (in 1902-1903) of the parallel relationship between cytological chromosome behaviour in meiosis and Mendel's laws. Guyer's suggestions were among the first, which attempted to relate the variation observed in the offspring in hybridisation studies by a coherent cytological chromosome mechanism to meiosis before the rediscovery of Mendel's principles. This suggested for the first time that the chromosome mechanism involved a conjugation of maternal and paternal chromosomes during the synapsis followed by a segregation of parental chromosomes in the final germ cells and a random union of the final germ cells in the fertilization. It shows that this early suggestion was similar to William Austin Cannon's later chromosome proposal attempting to explain Mendel's principles and had some influence on Walter Sutton's cytological suggestion explaining correctly the behaviour of Mendel's particle by 1903. |
Introduction
The role of cytological knowledge in the late 19th century in the setting up of the 'Sutton-Boveri chromosome hypothesis' proposed in the early 20th century has been fully debated. On one hand, historians have suggested that the rediscovery of Mendel's laws in 1900 gave a conceptual framework necessary to clarify the cytological findings of the decade of 1880-1890 and to lead to the acceptance of the link between chromosomes and Mendelian factors. According to Alice Baxter and John Farley, the cytologists of the late 19th century lacked a theory of hereditary transmission to explain the behaviour of the chromosomes in the process of meiosis (Baxter and Farley 1979). For Jean Gayon, the cytological findings in the years 1880-1890 were much too obscure to serve as a basis for an exact theory of heredity and probably delayed the construction of an experimental science of heredity based on hybridisation (Gayon 1993, 41-42).
On the other hand, historians have claimed that Mendelism would have been impossible without the cellular knowledge gained in the late 19th century. For Lindley Darden, the segregation concept noted by the cytologists in pollen and egg cells in hybrid was 'the key Mendelian discovery' (Darden 1991, 55). For Robert Olby, the cytological processes and the material particles related to Mendelian inheritance were already known in 1880-1890. Thus, he claimed, the cytologists would have probably expressed the Mendelian principles if the hybridists had not rediscovered them in 1900 (Olby 1966, 136). William Coleman, in an extensive historical study, has reported the progressive recognition of the role of the nucleus in heredity during the second half of the 19th century and its importance for the acceptance of the chromosomes as vehicles for the Mendelian factors (Coleman 1965). In reprinting important papers dealing with the study of inheritance in the late 19th century, Bruce Voeller has attempted to show the decisive contribution of cytology to the acceptance of the chromosome theory by 1903 through the progressive recognition that the chromosomes were the individual bearers of the hereditary information (Voeller 1968).
Despite this controversy, it is noted in the history of cytology and genetics that the similarity between the principles followed by Mendelian factors and the behaviour of chromosomes during meiosis was pointed out two years after the rediscovery of Mendel's law in the chromosome theory of heredity1 of Walter Sutton (Sutton 1903) and Theodor Boveri (Boveri 1904, 116-119). Before 1900, it is claimed, the attention of cytologists was primarily focused on the study of the chromosomes of pure-bred organisms. It was only after the rediscovery of Mendel's laws that they began to investigate germ-cell formation in hybrids during meiosis in order to define a physical basis for hereditary principles (Richmond 2001, 58). However, many of the studies focusing on the relation between heredity and cytological observations around 1900 have been based on German or British works. Except Sutton's studies (McKusick 1960), little attention has been given to American cytologists concerned by heredity studies during this period (Maienschein 1987; Paul and Kimmelman 1988). However, as Diane Paul and Barbara Kimmelman have stressed, studies concerning inheritance of characters in plants or animal were of extraordinary importance in United States at this time. For most
researchers, whether or not they wanted to improve selection or find theoretically a physical basis of heredity, breeding work was of central importance to understand the laws of heredity for practical or theoretical aims (Paul and Kimmelman 1988).In this paper, we will focus attention on the early work of the American cytologist Michael Frederic Guyer (1874-1959)2 well known to have contributed to the debate on the chromosome theory between 1906 and 1910 (Darden 1991, 91; Sapp 1987, 17). We shall claim that his doctoral dissertation Spermatogenesis of Normal and of Hybrid Pigeons (Guyer 1900a), published as early as 1900, was one of the first attempts to establish a parallel between the observed variation in offspring in crosses and the cytological chromosomes behaviour in the meiosis before the rediscovery of the Mendel's principles. We will show that this connection between observed hereditary traits across generations and chromosomes behaviour was strongly supported in Guyer's opinion by his assumption of chromosome individuality an his hypothesis of the maternal and paternal chromosomes association during the synapsis stage followed by a separation of the parental chromosomes in the final germ cells. It is historically3 accepted that the concept of maternal and paternal chromosome association was first expressed in the work of Thomas H. Montgomery later in 1901 (Montgomery 1901) and many historians consider this concept as a crucial step, which allowed cytologists to link the chromosomes and the Mendel's factors as early as 1902.4 In the light of some available published sources, we will finally discuss the reception of Guyer's hypothesis later by some of his contemporaries such as Walter Sutton and Austin Cannon, two American scientists who are considered to have suggested for the first time that the Mendelian ratios had their basis in the cytological phenomena of meiosis and fertilization
{1 Lindley Darden has reported on other theories called also 'chromosome theory of heredity': the claim in the late of the nineteenth century that the chromosomes were the bearers of the hereditary material and the theory of germplasm of August Weismann (Darden 1991, 81).
2 Michael F. Guyer studied zoology at the University of Chicago where he worked out his doctoral dissertation during 1897-1900 on pigeon spermatogenesis under professor Charles Otis Whitman. He became then professor at zoology at the University of Cincinnati where he continued his research on spermatogenesis of animal species and their hybrids (Noland 1972).
3 See Voeller (1968), Darden (1991), Baxter and Farley (1979).
4 The content of several papers reveals this early link between Mendelism and cytology: Wilson (1902), Cannon (1902a), Sutton (1902), Sutton (1903), T.H. Montgomery (1903) and Montgomery (1904).}
Historical background of the Sutton-Boveri chromosome theory
During the second half of the 19th century and the first years of the 20th, intensive exploration in the fields of cytology have led to the establishment of many of the fundamental observations underlying the origins of the Sutton-Boveri chromosome theory of 1903-1904. The increase of knowledge of the cytological details of fertilization conducted by Hertwig, Fol and Strasburger in the mid-1870s and the significance of the role of the nucleus as the bearer of the hereditary characteristics claimed by Strasburger, Hertwig, Weismann and Kolliker in 1884-1885 was established (Voeller 1968, 3). In reporting the cytological works - maturation, fertilization and division of the germ cells - in the 1890's in his monumental treatise
The Cell in Development and Inheritance in 1900, Edmund B.Wilson was able to demonstrate the central importance of the nucleus and its component, chromatin, as 'a leading factor in inheritance' (Wilson 1900, 359). During this same period, the cytological details of meiosis were closely examined but its interpretation remained controversial (Baxter and Farley 1979;Churchill 1970). The disagreement was about whether or not a qualitative reduction of the hereditary material occurred during the last steps of reduction division and on the permanence of the chromosome structure.For some scientists, led by Johannes Ruckert, the meiosis had to be interpreted in terms of Weismann's hypothesis of a qualitative reduction of the hereditary characters. In their opinion, the maturation division process was both quantitative and qualitative. Each chromosome tetrad observed before the first meiotic division consisted of two chromosomes joined end to end divided longitudinally. The first division was equatorial because it separated identical chromosomes along the longitudinal cleft in each tetrad. The second division was reductional as it separated the two chromosomes joined end to end along a transverse axis. The initial appearance of half the normal number of chromosomes in the first stages of meiosis was caused by a pseudoreduction of the chromosomes, implying that the usual somatic number of chromosomes was still present in the cell. The belief in the permanence and individuality of the chromosome during the cell division and the maturation of the germ cells was the strongest support for this theory to which Boveri might have accepted finally in 1904.5
{5 During the 1890s, Boveri's opinion on the meiosis was opposed to Ruckert's opinion. Each chromosome tetrad resulted, in fact, in two succeeding longitudinal divisions of one chromosome. Each resulting part from the division in the cells was thus equivalent and there was no qualitative reduction (Baxter and Farley 1979, 149-150). Some years later, Boveri had completely changed its mind in favour of Ruchert's theory after his crucial experiments on the multipolar mitoses in sear urchin eggs (Baxter and Farley 1979, 162-163).}In one another group, with cytologists such as Hertwig, Brauer and Guignard, the chromosomes were only transitory aggregating chromatin particles appearing from time to time. The first and second meiotic division were only quantitative and no qualitative reductions occurred. The final germ cells were equivalent in regard to hereditary properties. These scientists denied the permanent nature of chromosomes. As Alice Baxter and John Farley have stressed, the resolution of the controversy about the reduction division occurred when the uniqueness and the individuality of the chromosomes were clearly accepted once the correspondence between the chromosomes and the Mendelian factors was pointed out into the Mendelian scheme. A major step for the acceptance of the chromosome permanency throughout cellular division was the proposal in 1902 by the American biologist McClung that the 'accessory chromosome' discovered by the German zoologist Henking was related to sex determination (McClung 1902). It is also admitted among historians of genetics that the central question whether each chromosomes differed from the others in carrying a separate portion of hereditary determinants was also resolved by the work of Thomas Montgomery in 1901 (Montgomery 1901).
Grounded on chromosome morphology Montgomery concluded that the maternal chromosomes were paired only with paternal ones during meiosis. However, as Alice Baxter and John Farley noted, Montgomery's hypothesis of chromosome pairing was grounded on his theory of rejuvenation (Baxter and Farley 1979, 162). In his opinion, sexual reproduction was a mechanism of rejuvenation. The conjugation of the maternal and paternal chromosomes during the synapsis stage served to 'rejuvenate' the chromosomes and consequently the egg became rejuvenated by the fertilization with the sperm. It is in this cytological context briefly described above that Guyer's dissertation may be considered.
Guyer's dissertation Spermatogenesis of Normal and of Hybrid Pigeons
This paper containing 61 pages and 50 figures is a cytological study mainly reporting the description of the different phases of spermatogenesis in two species of pigeons ('common pigeon' and 'ring dove' species) and the hybrid form resulting from the crossing of these two species. Although it shows some figures, it mainly uses written descriptions.
In his introduction, the author presented his work as an attempt at the 'description of the various phases undergone by the germ cells in their transformation from spermatogonia to mature spermatozoa' (Guyer 1900a, 3). His choice of the pigeon for a cytological study was based on the lack of cytological studies concerning the sexual cell formation in this species,6 but his decision was also supported by the fact that his PhD supervisor Charles Otis Whitman was strongly engaged in research with these animals.7
{6 'So far, it seems that no investigations upon the spermatogenesis of the pigeon have been recorded. A few scattering observations have been made upon the spermatozoa, but the subject has been, in the main, untouched' (Guyer 1900a, 3).
7 Charles Otis Whitman (1842-1910) played an important influential role in American biology (Maienschein 1987, 81) and was perhaps one of the most famous embryologists of his time (about Whitman's biography, see Mayr (1976)). His research focused on the heredity and the variation of the pigeons in breeding. As an hybridist, he performed crosses of some 200 domestic varieties and 40 wild species. His results were recorded in a posthumous work (Riddle 1919).}
However, in focusing a part of his PhD study on the spermatogenesis of the germ cells in hybrid forms, the author was convinced that his work on hybrid forms might lead to wider conclusions on the phenomenon of heredity:
|
It is a remarkable fact that no attempt has been made so far to investigate carefully the spermatogenesis or ovogenesis of hybrid forms. In all the mass of literature discussing or touching upon hybridism, so far as I have been able to ascertain, there has been in no instance an approach to a thorough study of the germ cells. Yet almost every writer states that through the study of hybrids, we have perhaps the best opportunity for gaining a clew to many of the most vital points in the great problem of heredity (Guyer 1900a, 34). |
Through his description of the different cellular stages during the meiosis in the pigeon, the author was fully concerned by the problem dealing with interpretation of the reduction division8 and the synapsis stage9 as other scientists of his time.10
8 The term 'reduction division' was used since 1887 in the descriptions of spermatogenesis and oogenesis to mean a qualitative reduction in characters (Baxter and Farley 1979, 148).
9 'As to the meaning of synapsis, we seem to be in almost total ignorance' (Guyer 1900a, 17). The synapsis stage is characterized by the joining of the homologous chromosomes (maternal chromosomes with paternal ones) before the first division of meiosis.
10 See the chapter 'Historical background of the Sutton-Boveri chromosome theory' presented above.
In observing that the primary spermatocyte in normal (non-crossed) pigeons presented half the original number of chromosomes in the meiotic synapsis (Guyer 1900a, 17), Guyer's main aim in his dissertation was to understand how this process of chromosome reduction occurred during the synapsis stage. However, because he had some difficulties to follow correctly the chromosome behaviour during meiosis in the normal pigeons, Guyer decided to focus his observations on spermatocytes of hybrid forms of pigeons. He had observed that the chromosomes in these spermatocytes were easier to follow because they were not fusing together in the synapsis stage. In hybrids from very distinct species, this non-fusion of the chromosome was especially clear under the microscope, leading to better observations on the behaviour of the chromosomes during the synapsis:
|
In the spermatocytes of normal pigeons there is no satisfactory evidence other than the remarkable decrease in the number of chromosomes to indicate the exact nature of the division but in hybrids, owing to certain exaggerations which frequently occur, especially in hybrids from very distinct species, some very significant facts come to light (Guyer 1900a, 25). |
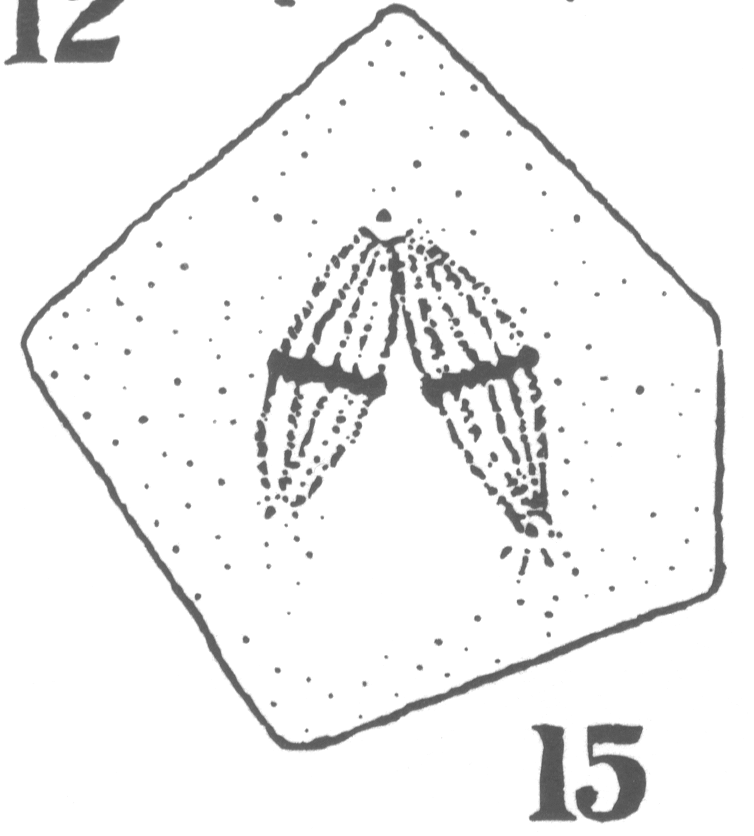
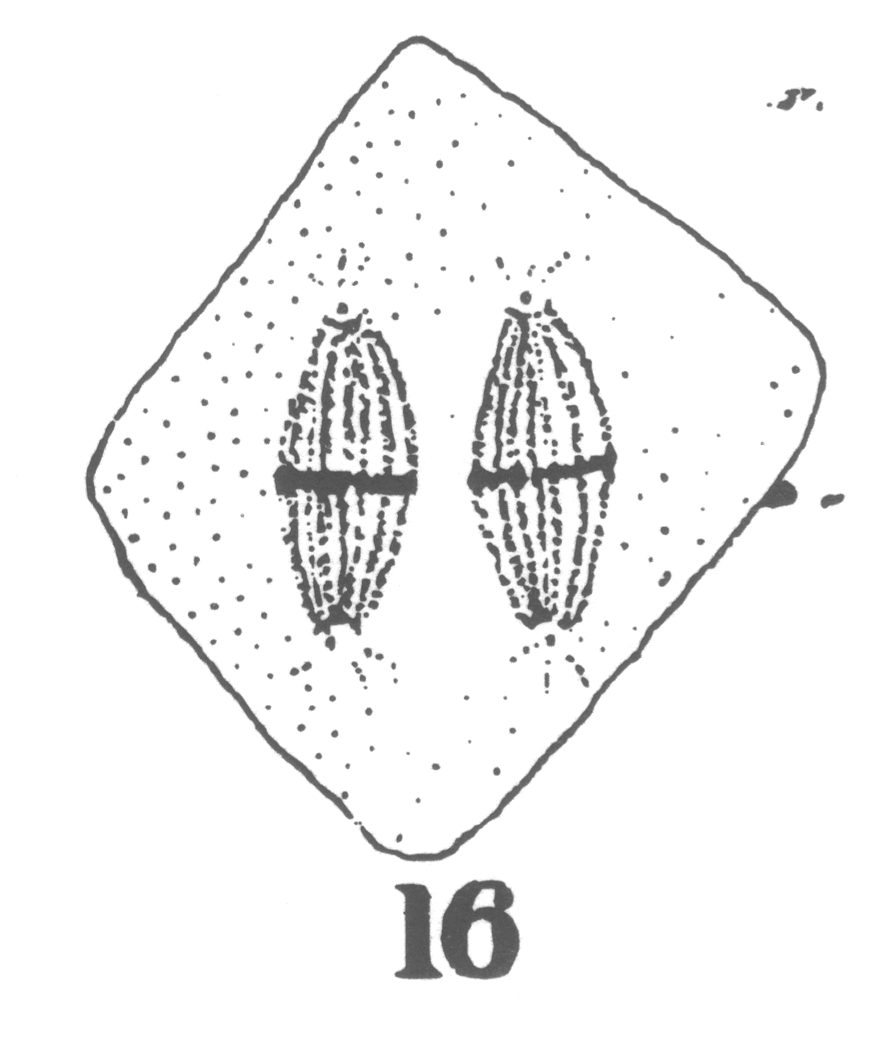
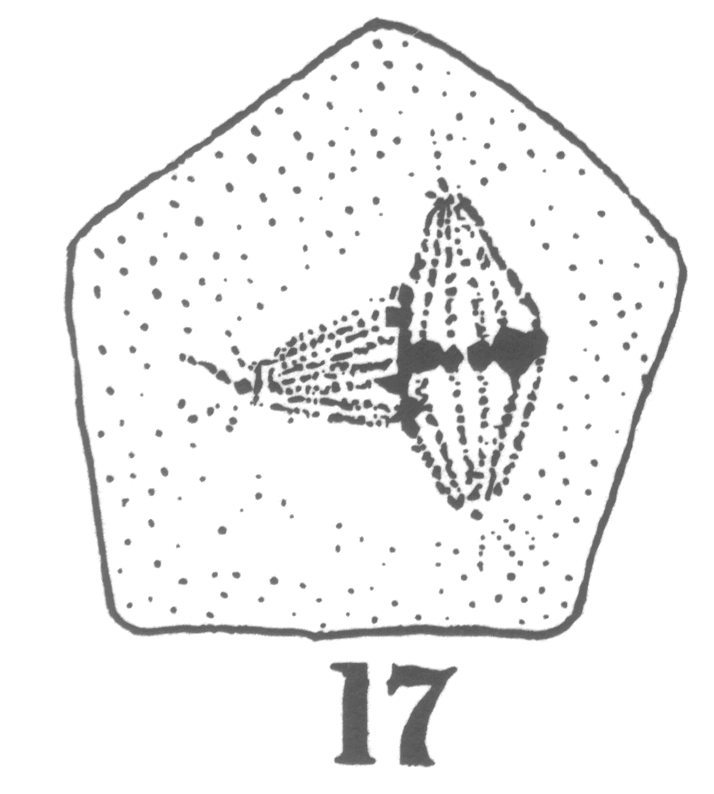
Guyer reported from his observations that in the primary spermatocytes of hybrid forms the 'pairing' necessary to the formation of bivalent chromosomes characteristic of the synapsis stage 'comes about with more or less difficulty or not at all' compared to spermatocytes in the normal pigeons (Guyer 1900a, 26). The chromosomes remained in two groups generally on two separate spindles until both groups were finally separated during the succeeding division (Fig. 1) (Guyer 1900a, 38-39). Each final cell formed at the end of the spermatogenesis in hybrids thus contained exclusively chromatin originating from one of the spindles (Guyer 1900a, 40).
 |
 |
 |
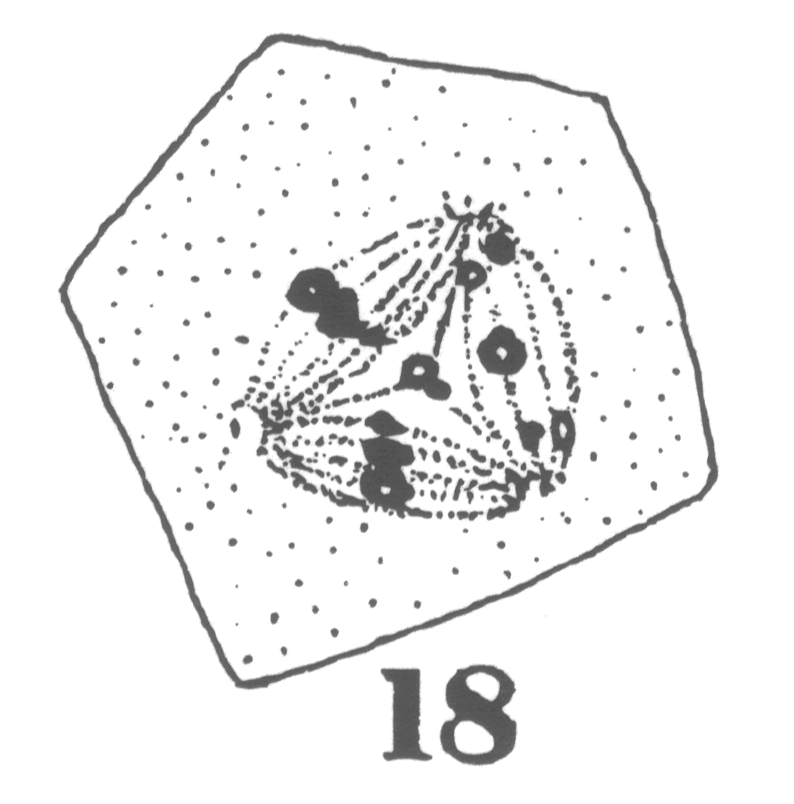 |
Fig. 1 - Guyer 1902, 10. The linear sequence of irregularities in division of the primary spermatocytes (showing the multipolar spindles) in hybrid pigeons.
Guyer inferred that the chromatin present in each final group that he observed probably came from each parent species of the hybrid. This suggestion was supported by the cytological observation of the abnormal formation of the spindle in the hybrid cells11 but was also by the previous conclusions of hybridisers who had observed in the somatic cells of hybrids 'the influence of two distinct tendencies' and had attributed this to an intermediate type between the two parent types (Guyer 1900a, 46).12
{11 'It [the irregularity in the formation of the spindle] simply means, -- that in all probabilities there is an incompatibility between the chromosomes from the two different species' (Guyer 1900a, 39).
12 Guyer referred here to a study of J. Murehead Macfarlane (Murehead Macfarlane 1891) which had concluded that hybrid forms were intermediate between the two parents after observations of many plant crossbreeds.}
Thus, he inferred that if the chromosomes of each of the two parents in a sexual cell of a hybrid kept their own individuality without joining together, the following cellular division might consequently segregate the maternal and paternal chromosomes in separate germ cells (Guyer 1900a, 55). On a priori grounds, this consideration led him to general considerations concerning the chromosome behaviour in the second division of the meiosis. If these irregularities, which were observed in the meiosis in the hybrid forms crossed from very different species reflected what really occurs during the meiosis in the sexual cells of the offspring from ordinary cross (Guyer 1900a, 49),13 the synapsis stage observed in the meiosis of ordinary crosses would result from an 'union' (Guyer 1900a, 49) or a 'conjugation' (Guyer 1900a, 17) of the maternal and paternal chromosomes.
{13 Guyer stressed the similarity of the meiotic process between the germ cells of hybrids from very different form and the germ cells of common hybrids: 'One is forced to the conclusion-- that the double-spindled and multipolar types of cells which occur in hybrids between very different forms are but exaggerated images of a tendency which exists in the primary spermatocytes of normal appearance, which are to be found in all hybrids' (Guyer 1900a, 55-56).} A separation of the maternal and paternal chromosomes (called here plasmas) would then occur in different germ cells during the last stage of the genesis of the sexual cells:There is but one ultimate conclusion, then, namely, that the irregularity in division of the primary spermatocytes which appears in hybrids between very different species, is but an index to what occurs in ordinary crosses. In the latter, instead of separate spindles and non-fusion of chromosomes, a true union occurs, but the bivalent chromosomes ultimately divide in such a way that the respective plasmas occupy in different cells. There is a separation of the paternal and the maternal chromosomes which had fused during synapsis (Guyer 1900a, 49). |
Following this idea, he concluded that the final germ cells after this segregation of the maternal and paternal chromosomes might be considered 'pure' because they would contain 'qualities of only one species' (Guyer 1900a, 55), or in other words, because the final germ cells might include in his opinion either all the maternal chromosomes or all the paternal chromosomes. This hypothesis of parental chromosome segregation in the final germ cells implied naturally the belief that the chromosomes were entities persisting throughout the whole meiotic process:14
{14 His acceptance of the chromosome persistence was especially based on the fact that the sex chromosome (he called 'accessory chromosome' of McClung) was visible all the time during the meiosis in his observations (Guyer 1900a, 13).}With regard to the question of the persistence of chromosomes, the evidence is becoming stronger every day that these elements do retain their individuality. In addition to the well known observation of Boveri on the early stages of Ascaris, there are numerous other facts which point to the same conclusion. Ruckert for instance, in his study upon the fertilization of cyclops, was able to follow the maternal and paternal chromosomes very distinctly in cleavage. -- Again, to cite one more of the rapidly multiplying examples, perhaps the most brilliant confirmation is met with in the work of Herla and of Zoja. Both observers have shown that in the hybrid fertilization of Ascaris, -- the three chromosomes thus brought together retain their individuality and reappear at each cleavage, at least to the twelve-cell stage (Guyer 1900a, 49-50). |
However, his suggestions concerning the maternal and paternal chromosome union in the synapsis stage and the consecutive separation of the maternal and paternal chromosomes in the final germ cells were also grounded on his observations on the colour trait of the doves observed in the successive generations in crossing:
In chromatin we have a substance which from all we know of its nature and actions, seems to be intimately bound up in the phenomena of inheritance. It is reasonable to suppose, therefore, that it constitutes at least a part of the material basis for the variation in the germ. The questions then arises as to whether there is any correlation between the distribution of chromatin as seen in the germ-cells of hybrids, and the marked variability which characterizes the offspring of fertile hybrids (Guyer 1900a, 46-47). |
Since the beginning of his dissertation in 1897, Guyer carried out observations on successive crossing of the two studied pigeons species as one of the earlier reports on his PhD study shows (Guyer 1899). The influence of his supervisor Whitman was of first importance in such work, as Whitman came from a long tradition of hybridists and was directly concerned with crossing studies.15
{15 See footnote 7. As a contemporary of Hugo de Vries, Whitman has been very impressed by his theory about the 'unit-characters' writing in 1897: 'It is to a comparative and experimental analysis of specific characters that we must look for a knowledge of the phenomena of heredity and variation' (Riddle 1919, vol. I, iv). However, Whitman became over time more doubtful that the ideas of de Vries could explain heredity fully, but he recognized the remarkable parallelism carried out between the cytology findings and the Mendelian ratios, and he wrote in 1909: 'The idea of unit-characters, however, as distinct elements that can be removed or introduced bodily into the germ does not appeal to me as removing difficulties, but rather as hiding them; -- I am strongly persuaded that his hypothesis of unit-characters fails as a guide to the interpretation of the species and its characters. It is true that a great amount of work on Mendelians heredity seems strongly to support the unit-character hypothesis, and that cytology offers some further support' (Riddle 1919, vol. I, 5).}
From the crossing of his two species of pigeon respectively brown and white in the plumage, Guyer reported that the hybrid offspring was brown in the second generation. The crossing between the hybrid offspring gave third generation offspring, which presented generally with a 'reversion' of the character16 and thus 'a return to the original colors of the grand parents' (Guyer 1900a, 35).
[16 The hybridists were familiar with the so called 'reversion' of the character which was the observed phenomenon of the reappearance of ancient characters in the next generations. In early 1897 Whitman had itself previously established this phenomenon in crossing pigeons: 'From my own summer's work with 3 pairs of parents -- .The point of chief interest in these results is this: In all these pairs, every one of the parents being brown, we get quite a large proportion of white young birds. In other words, the white color of the parent bird shows not in the second, but in the third generation' (Riddle 1919, vol. II, 144).{The offspring from this generation were respectively white and brown but sometimes both of the young were brown or less frequently white, as he explained:
So far as the writer has carried his experiments, the indications are that on the whole there are more brown than white birds in the third generation, and this points to the conclusion that in the brown birds we may have both intermediate forms like the hybrids of the second generation and forms which have reverted to the brown grandparent, as the white doves have seemingly returned to the white grandparent (Guyer 1900a, 36). |
However, Guyer stressed in his conclusions that the white birds he had observed in the third generation were not all due to reversion to the white grandparent, but might be the results of intermediate forms. In that case, these intermediate forms might be white just because sometimes one of the brown parents sometimes showed a lighter shade of brown, leading to offspring showing different 'degrees of intermediacy' of the brown colour but remaining intermediate in other characters (Guyer 1900a, 36). Concluding his observations on the crossing of the doves, Guyer inferred that a greater number of intermediate forms than reverted forms might occur in the third generation. His assertion was grounded on cytological considerations:
As we shall see in the conclusions from the study of the germ cells of hybrids, there are certain phenomena in the germ cells which apparently afford us a definite physical basis for the production of intermediate forms and for returns to pure ancestral species. From this basis there must necessarily be a greater number of intermediate forms in the offspring of hybrids than there are reversions to the respective ancestral species (Guyer 1900a, 36). |
It might be stressed two important points.
First, Guyer did not perceive the independence of the characters in segregation, one of the central aspects of Mendelian genetics. He clearly believed that the behaviour of the colour trait among generations was strongly related to the behaviour of other traits. That means, in other words, that he interpreted the reversion of the offspring of the third generation to the grandparent types for the whole parental trait, and not just in one trait in particular. The colour trait was just used in his crosses because it was a representative characteristic of the species crossed, but he did not notice that this trait might be independent of the other species traits in their behaviour.
Secondly, he did not express in his work the percentage of white and brown offspring he had in the third generation, as the doves was difficult to use as material for a hybridisation study to get definitive statistical results.17 {17 'Inasmuch as doves produce two young at a brood, it would require a long period of time to secure numbers sufficient to arrive at satisfactory conclusions regarding the percentage of actual reversion to the original species' (Guyer 1900a, 36).} However, in our opinion, he had sensed the need to explain phenotypic variation in hybrid offspring in using statistical methodology. For instance, in discussing if some irregular distribution of chromatin in the meiosis might explain the observed case of reversion to one parent only, Guyer stressed how he had observed that the frequencies of such hypothetical distribution might be 'by far too few to equal the percentage of reversions' (Guyer 1900a, 49) he had obtained from his crossing experiments. It seems obvious that the influence of Charles Benedict Davenport who was an assistant of Whitman in Chicago at this time had some influence on this statistical methodology,18 although no part of the Spermatogenesis mentions the name of Davenport. {18 Charles Benedict Davenport (1866-1944) was a pioneer in the statistical studies of populations since 1890 and published during his life several quantitative studies on the use of biometric methods in the United States such as his manual Statistical Methods with Special Reference to Biological Variation (1899) (about Davenport' biography, see Shor 1971). Following Paul and Kimmelman (1988), Davenport was one of the first to introduce Mendelism in America (see Davenport 1901). He became assistant professor at the University of Chicago in 1899 in the department of Charles Otis Whitman.}
In relating his results from hybridisation studies to his cytological observations, Guyer stressed how both processes were strongly interdependent and led to gametes bearing different hereditary qualities through a separation of the paternal and maternal chromosomes (that he referred to plasmas) resulting in a qualitative reduction of the characters:
| To explain reversion and variation in the offspring of fertile hybrids, we are forced to the conclusion that a segregation of the paternal and maternal plasmas occurs in the germ cells, and the only chance for such a segregation, if the cells are of the normal type, is through the reduction division (Guyer 1900a, 26). |
This qualitative reduction occurred mechanically through a pseudoreduction in Johannes Ruckert's sense:
| Synapsis occurs in the primary spermatocytes through which a pseudo-reduction of the chromatin takes place. In the division of the spermatocyte only eight chromosomes appear, but they are in the form of heavy rings or vesicles and are evidently bivalent (Guyer 1900a, 33). |
Finally, in pooling together his conclusions drawn from cytological observations and from hybridisation studies, Guyer suggested a mechanism of chromosome segregation attempting to explain the physical variation and the phenomenon of reversion observed in his offspring in the third generation:
| There seems to be no other interpretation, indeed, than that in the many normal mitoses [meiosis] of the bivalent chromosomes which occur, the chromatin of the father and of the mother is set apart so that the ultimate germ-cells are what might be termed 'pure' cells; that is, a given egg or sperm-cell contains exclusively or at least predominantly qualities from one parent. The offspring from fertile hybrids of the same parentage might then be similar to the mixed type of the original hybrid, or revert to one of the grandparent types, dependent upon the chances of the various cells for union at fertilization. If a spermatozoon and an egg containing characteristics of the same species unite, then the reversion will be to that species; if a sperm-cell containing the characteristics of one species happens to unite with an ovum containing characteristics of the other species, then the offspring will be of mixed type again. By the law of probability the latter will be the more prevalent occurrence, because there are four combinations possible, and two of the four would result in the production of mixed offspring, while only one combination could result in a return to one of the ancestral species (Guyer 1900a, 47-48). |
In our opinion, this above suggestion represents an important step in the history of genetics. For one of the first time, the three-fold ratio 1:2:1 of offspring in the third generation was explained by a coherent cytological mechanism involving chromosome segregation in the final germ cells in meiosis and random union of these chromosomes in fertilization. Such an idea was expressed later by Correns, one of the discoverers of Mendel's law, through the behaviour of his hypothetical 'anlagen', which when joined at random, might give in the third generation one of the parental types of the first generation or the hybrid type of the second generation (Corcos and Monaghan 1997, 404). It might be stressed that this 1:2:1 ratio was available in Guyer's mind for the whole parental set of characters and not only for a particular species trait, because he did not perceive at that time that the parental characters might be not associated together during segregation. Because Guyer supported his chromosome mechanism and his idea of the effect of the germ-cell 'purity' on the behaviour of the colour trait of the doves through the generations, he was thus able to express correctly this ratio for the results of crosses in which the parents differ only by a single character. For unknown reasons, he did not study the behaviour of the other traits in doves in the crossing experiments in his dissertation. Despite that he based his suggestion on a single trait behaviour, we stress that his proposal may probably be one of the earliest coherent hypothesis which attempted to establish a connection between the cytological phenomena and hereditary phenomenon of reversion without the knowledge of the theoretical principles of Mendelian inheritance. It is of particular interest to note that the misunderstood 'reversion' of the characters observed in the third generation of hybrids was explained according to this suggestion by the characters of the parents of the first generation only and not by ancestral characters as it was the case with Galton's laws.19 {19 In Galton's theory, each ancestral parent contributed a proportional part of the total heritage of the descendant: the two parents contribute one-half, the four grand parents one quarter, the eight grand parents one-eighth and so on. The inheritance was mediated through elements which could be expressed only in subsequent generations, thus explaining the phenomenon of reversion (Bulmer 1999).}
Furthermore, it seems obvious that his idea of conjugation of maternal and paternal chromosomes during synapsis was similar to the suggestion expressed by Montgomery in 1901 in writing that 'the synapsis stage -- may be the stage of the conjugation of the chromosomes' (Montgomery 1901, 223). However, in his paper Montgomery did not quote Guyer's work. Interestingly, Guyer did not base his hypothesis of maternal-paternal conjugation of the chromosomes during the synapsis stage on the rejuvenation theory as Montgomery did to support his own hypothesis of pairing. It was supported by a pure cytological model of chromosome behaviour explaining in a coherent way the observed variation of the offspring in the third generation.
Finally, we stress how this earlier proposal of Guyer of maternal-paternal chromosome union and its consecutive segregation may recall the later suggestion of Walter Sutton 'that the association of paternal and maternal chromosomes in pairs and their subsequent separation during the reducing division -- may constitute the physical basis of the Mendelian law of heredity' (Sutton 1902, 39).
However, Guyer suggested some other cytological hypothesis to explain the variations he had observed among the offspring in the third generation of his crosses. Without the knowledge of the Mendelian principles, he did not point out the universal nature of his hypothesis concerning the maternal and paternal chromosome segregation in the final germ cells and their random union in fertilization. In fact, Guyer did not think that this chromosome mechanism could account for all the phenotype variations observed in the crosses. His main aim was to bring cytological suggestions for explaining highly diversified gametes, which through union might explain the 'marked variability' (Guyer 1900a, 47) in the plumage colour of the offspring resulting from his crossing experiment.
Thus, many of his other cytological suggestions in his dissertation were to be criticized later by his contemporaries in the light of the Mendelian inheritance principles.20 For instance, Guyer proposed that the chromosomes of the secondary spermatocytes could also divide in such a way that both maternal and paternal chromosomes would be included in the same cell (Guyer 1900a, 27). However, this suggestion concerned only the spermatogenesis in the normal forms of pigeons. Furthermore, he claimed that inequality in the distribution of chromatin in the germ cells might also sometimes occur,21 although he remained convinced that greater part of the variation recorded among the offspring of hybrids was explained by a separation of maternal and paternal chromosomes in 'pure' germ cells and the random union of two 'pure' germ cells (Guyer 1900a, 48-49).
Finally, he was also convinced of the role of cytoplasm in the hereditary transmission.22 In his opinion, the cytoplasm brought a 'general substratum' providing the general characters of the species on which were superimposed the specific characters of the individual conveyed by the chromatin (Guyer 1900a, 51). This belief concerning the importance of the cytoplasm in transmitting hereditary determinant was grounded on the experiments of Hans Driesch having shown with Thomas Hunt Morgan in 1895 that the removal of cytoplasm from the uncleaved ctenophore egg might give defective embryos (Guyer 1900a, 53). This idea about the role played by the cytoplasm in the hereditary process became a major recurring theme in his works after 1903.23
{20 See below the chapter entitled 'Reception of Guyer's hypothesis by the contemporaries'
21 'From the fact that in cases of apparently complete return to one parent type, characteristics of the other parent may nevertheless crop out from time to time in succeeding generations-- The occasional inequalities in the division of individual chromosomes -- may account for this fact' (Guyer 1900a, 48).
22 'There seems to be no sufficient reason for not regarding the cytoplasm likewise as an important
factor' (Guyer 1900a, 51).
23 See for instance Guyer (1911).}
Guyer's hypothesis and the rediscovery of Mendel's principles
Although the thesis publication was dated from 10th May 1900 one month after the French publication of Hugo de Vries' report of the Mendel's laws rediscovery (Vries 1900), it seems obvious that Guyer was not aware of the Mendelian rediscovery in presenting his concept of chromosome segregation and 'pure' germ cells. First, the thesis was accepted by the University of Chicago in March 1900,24 indicating that the paper has been prepared at an earlier time. {24 This was indicated by a summary of the thesis published later in the Bulletin of the University of Cincinnati (Guyer 1902, 3)}. Second, in two successive later papers reporting his earlier ideas of the relation between chromosome behaviours in the germ cells of the hybrid and the reversion phenomena of the offspring in the third generation (Guyer 1903a; Guyer 1907), Guyer claimed full ignorance at this time of the Mendelian principles.25 {25 'In studying the germ cells of hybrid pigeons some years ago (1897-1900), the writer observed certain phenomena which led him to much the same opinion regarding the separation of qualities in germ cells as that expressed by Mendel, although at the time the results of Mendel were wholly unknown to him' (Guyer 1903a, 490); 'Although I had elaborated the idea of the relation between the chromosomal phenomena of hybrids and their progeny, in my doctoral thesis of 1900, the published statement of the details did come out until November, 1902, but these details show that I had hit upon practically the Mendelian ideas' (Guyer 1907, 1008).} Third, in a short paper published on February 16th 1900, before Mendel's rediscovery, Guyer himself reported one of the key ideas of his PhD thesis was that a maternal and paternal chromosome segregation might occur in separate germ cells:
| --it is evident that after some division, some of the new cells will have chromatin from only one of the original parent species and some, from the other. Some of the spermatozoa, therefore, will bear chromatin from one only of these species. It is a well-known fact that the offspring of hybrids are extremely variable, a portion of these variations being usually in the form of reversions to one or the other of the parent species. The possibility presents itself then, that this reversion may be due to the persistence of the chromatin of only one species in one or both of the germ cells (Guyer 1900b). |
His ignorance of the rediscovery of Mendel's law at the time of his doctoral work is strongly confirmed in his paper on May 1903. In the light of the Mendelian principle of the independent segregation of the characters, Guyer corrected his previous statement that no intermixture of paternal and maternal chromosomes occurred in the final gamete:
| -- It does not necessary follow that in the ensuing division all of the characters of one parent will be set apart in a separate cell-- If such a separation were actually to occur, then the offspring which returns to a grandparent type must revert in not only one given character, but in all characters; that is, the reversion would be complete. The Mendelian law, however, confines itself to a given character, and if any other character is chosen, although it will follow the same law, it does so without any reference to the first character, so that offspring may be pure with respect to a particular character, yet also possess other characters of a mixed nature, or even pure characters of the other parent (Guyer 1903a, 491). |
It seems obvious that he had understood that his earlier suggestions only explained crosses in which the parents differ by a singlecharacter. Thus, in the beginning of 1903, in stressing that paternal and maternal chromosomes should be not considered as an inseparable group in the segregation process, he had drawn the same conclusions as Sutton in his paper 'the Chromosomes in Heredity' (Sutton 1903) in assuming that an independent segregation of the paternal and maternal chromosomes in the final gamete during gametogenesis might fully explain the Mendelian theory.26 {26 Although Guyer's article was published in the beginning of May 1903, it was the written account of a conference held on 18th April in the same time of the publication of Sutton's paper (April 1903).}
Reception of Guyer's hypothesis by the contemporaries
A. Baxter and J. Farley stressed that the correspondence between the behaviour of the chromosomes during meiosis and Mendel's principles was not been made immediately by cytologists in 1900, but that it took a while until Montgomery's ideas of chromosome pairing during synapsis was fully accepted (Baxter and Farley 1979, 165-166). However, in their assertion these authors did not take into consideration Guyer's work. In his paper of 1907, Guyer claimed how his early ideas linking the chromosomes to inheritance facts developed in his thesis by 1900 were similar to later suggestions of cytologists involved in the physical explanation of Mendelian precepts:
| Various hybridists, judging from the external visible characters of hybrids, came to the conclusion that in the germ cells of hybrids there must be a separation of parental qualities so that with respect to a given quality, half of the germ cells returned to the maternal, half to the paternal types. Conversely, I had in the meantime from studying the germ cells of hybrid doves and pigeons, called attention to the fact that there is apparently a segregation of maternal and paternal chromosomes at the reduction period which, if the chromosomes bore hereditary qualities, would lead to the establishment of pure germ cells, and thus afford a possible explanation of how the offspring of hybrids come to show returns to grandparent types. -- Later Montgomery (1901) and Sutton (1902), working on non-hybrid forms, which have chromosomes of varying size, presented striking evidence of the fact that in early germ cells there are pairs of homologous chromosomes, one of paternal and one of maternal origin, and that these homologous chromosomes unite in synapsis. The ensuing reduction division simply brings about their separation and segregation in different cells. Although I had elaborated the idea of the relation between the chromosomal phenomena of hybrids and the reversion of their progeny, in my doctoral thesis of 1900, the published statement of the details did not come out until November, 1902, but these details show that I had hit upon practically the Mendelian idea. Very soon after the appearance of my paper in 1902, Cannon, Wilson and Sutton published conclusions of a very similar nature, endeavouring to offer a cytological interpretation of the Mendelian principles, which had their renaissance in the meantime. -- In this way it became established that there is a mechanism in germ cells, which, assuming that the chromosomes are concerned in heredity, could possibly lead to a segregation of paternal and maternal qualities (Guyer 1907, 1007-1008). |
In our view, it is obvious that Guyer's earlier suggestions were probably taken into account by the whole scientific community only at the end 1902, when a summary of his dissertation was published with the title Hybridism and the Germ Cell in the Bulletin of the University of Cincinnati (Guyer 1902). A reprinting of the whole thesis followed in the next issue of the Bulletin of the University of Cincinnati (Guyer 1903b). Many scientists responded enthusiastically to Guyer's idea on the chromatin distribution in the germ cells and cited Guyer's suggestions with approval in their own work (see Table 1).
| Name | Title | Published |
| Guyer, M. F. | Spermatogenesis of Normal and Hybrid Pigeon | May 1900 |
| Montgomery, T. | A Study of the Chromosomes of the Germ Cells of Metazoa | 1901 |
| Bateson, W. & Saunders, E. R. | Reports to the Evolution Committee. Report 1 | 1902 |
| Cannon, W. A. | Some Cytological Aspects of Hybrids | (held in 1902) |
| Guyer, M. F. | Hybridism and the Germ Cell | November 1902 |
| Cannon, W. A. | A Cytological Basis for the Mendelian Laws | December 1902 |
| Sutton, W. | On the Morphology of the Chromosome Group in Brachystola Magna | December 1902 |
| Wilson, E. B. | Mendel's Principles of Heredity and the Maturation of the Germ Cells | December 1902 |
| Guyer, M. F. | Spermatogenesis of Normal and Hybrid Pigeon (reprint) | 1903 |
| Cannon, W. A. | Studies in Plant Hybrids: The Spermatogenesis of Hybrid Cotton | March 1903 |
| Sutton, W. | The Chromosomes in Heredity | April 1903 |
| Guyer, M. F. | The Germ-Cell and the Results of Mendel | May 1903 |
| Boveri, T. | Ergebnisse Uber die Konstitution der Chromatischen Substanz des Zellkerns | 1904 |
Table 1 - Title and publication dates of Guyer's papers and articles citing Guyer's work.
For instance, William Bateson, one of Mendelism's most ardent champions although he was strongly opposed to the concept of material units to explain heredity (Darden 1977), the botanist William Cannon and the cytologist Walter Sutton quoted Guyer's conclusions. Cannon and Sutton are well known to have linked Mendelian genetics
and chromosome behaviour (Cannon 1902; Sutton 1902).In reporting Guyer's hypothesis, Cannon and Bateson both noted the link that Guyer drew between the chromosome behaviour and the observed phenomenon of 'reversion' in the third generation of hybrid offspring, stressing how these suggestions had been drawn in ignorance of the Mendelian rediscovery.27 {27 'This conclusion appears to have been drawn before the publication of the Mendelian law' (Cannon 1903, 152); 'Guyer suggests, through apparently in ignorance of Mendel's work-- ' (Bateson and Saunders 1902, 149).} Because he was more concerned with hybrid studies, Cannon was strongly impressed by the fact that Guyer's suggestion of chromosome behaviour during the meiosis could account for the Mendelian theory:
| If the form of maturation mitosis [meiosis] just described [by Guyer] occurs generally in hybrids, and further if the chromatin is separated and distributed as postulated by Guyer, we should evidently not have to go further in order to discover the kind of mitosis that would produce in hybrids of the Pisum-type 'pure' germ-cells - the important part of the Mendelian theory (Cannon 1903, 152). |
Stressing how the idea of chromosome pairing was important to support the connection between the Mendelian conception and chromosome behaviour, Cannon cited Montgomery and Guyer as the leaders of such an important suggestion (Cannon 1903, 154). More significant, in claiming that a separation of maternal and paternal chromosomes in the germ cells and their random union might explain the Mendelian ratio, Cannon noticed that Sutton's paper On the Morphology of the Chromosome Group in Brachystola magna and Guyer's paper Hybridism and the Germ Cell led to the same conclusions (Cannon 1903, 155). However, it seems obvious that Cannon had in mind Guyer's hypothesis of chromosome segregation expressed in his dissertation at an earlier time. In comparing Guyer's PhD dissertation of 1900 and Guyer's paper Hybridism and the Germ Cell of 1902, we can assert that the results expressed in these two papers are fully similar. There is no doubt that Guyer's ideas by 1900 had a strong influence on Cannon's hypothesis of chromosome segregation. In his famous paper A Cytological Basis for the Mendelian Laws by late 1902, claiming that a splitting of the chromosomes occurs during the meiosis so that each gamete would receive only paternal or maternal chromosomes and not both together28 (Cannon 1902, 660), Cannon was aware of Guyer's hypothesis of 1900. {28 Cannon believed that no intermixture of maternal and paternal chromosomes could occur in the final germ cells, which, of course, is not accepted today.} In a previous brief paper discussing the possible relation between experimental andcytological study of hybrids, Cannon quoted fully Guyer's suggestions about the relationship he had seen between parental chromosome separation in the germ cells and the variation observed in the hybrids (Cannon 1904). Thus, it can be concluded that Cannon's suggestions in 1902 and Guyer's hypothesis in 1900 were similar, and the main difference was that Cannon stressed the universal nature of the process in the light of the Mendelian principles:
| The chromosomes derived from the father and the mother unite in
synapsis and
separate in the metaphase of one of the maturation divisions, -- so
that the
chromatin is distributed in such a way that two of the cells receive
pure paternal,
and two cells pure maternal chromosomes, and no cells receive
chromosomes
from both the father and the mother. In this manner, it has been
demonstrated
that pure races of animals may, and normally do, organize sex cells
of pure descent.
Now since such is shown to be the case in pure races of animals, I
suggest that
the sex cells of fertile animal hybrids are formed in a similar way,
and thus we
may have in the animals a cytological basis for variation in accord
with the
Mendelian conception. And I further suggest that this is the case in
plants as well (Cannon 1902, 660-661). |
It is however Sutton who was to report the most on Guyer's suggestions in his fundamental paper of 1903 The Chromosomes in Heredity (Sutton 1903, 250-251). In his postscript, Sutton recognized the importance of Guyer's suggestions for the cytology, in quoting Guyer's paper Hybridism and the Germ-Cell of 1902:
| This investigator also has applied conclusions from cytological data to the explanation of certain phenomena of heredity, and his comparative observations on the spermatogenesis of fertile and infertile hybrids are an important contribution to the cytological study of the subject (Sutton 1903, 250). |
As did Cannon and Bateson, Sutton recognized how Guyer was familiar with the Mendelian principles of reversion in his work:
| No mention is made in the paper of Mendel's law but in considering the inbred pigeons hybrids from which his material was obtained, the author expresses his familiarity with manifestations of the Mendelian principle by the statement that 'in the third generation there is generally a return to the original colors of the grandparents' (Sutton 1903, 250). |
It is well known that Sutton's explanation of Mendel's law was essential to connect correctly the finding of cytology with the facts of heredity. In claiming that an independent assortment of the parental chromosomes occurs in the final gamete and fully agrees with Mendel's theory, Sutton corrected the previous conclusions drawn by Guyer and Cannon, which assumed that a complete separation of the paternal and maternal chromosomes might occur in the final germ cells (Sutton 1903, 250). However, because he had taken into account only the paper of Guyer of 1902, Sutton assumed wrongly that Guyer had expressed the same concept of chromosome segregation as Cannon at a later date:
| In assuming that there is a 'segregation of maternal and paternal chromosomes into separate cells, which may be considered 'pure' germ-cells containing qualities of only species'--, he repeats the error of Cannon-- (Sutton 1903, 250). |
Sutton stressed how Guyer's hypothesis of chromosome segregation in germ cells and a random union of two germ cells in fertilization could fully explain the result of Mendel's experiments, provided that his segregation explanation be not 'erroneously applied (and without cytological grounds) to all the characters or chromosomes instead of to individuals' (Sutton 1903, 250).
Go to: Guyer's 1900 thesis
(Click Here)
Go to:
Main Guyer Page
(Click Here) Return to Evolution Index
(Click Here) Return to HomePage
As McKusick has reported, there is no question of the originality of
the Sutton's idea concerning the independent segregation of
chromosomes to explain Mendel's principles (McKusick 1960, 490)
but it seems clear that the earlier cytological suggestion of Guyer,
which was made before the rediscovery of Mendel's laws was a crucial
step for Sutton in his setting up of the chromosome theory in 1903.
Concerning the establishment of the chromosome theory in Sutton's
mind, Edmund Beecher Wilson, one of the most important cytologist
experts working at Columbia University and a supervisor of Sutton,
wrote some few years later:
Only a year or two before Sutton, the work of Guyer, Montgomery and
Boveri
had led them almost to the verge of such an explanation. Sutton
however, was
the first clearly to perceive and make it known-- (McKusick 1960,
490).
Interestingly, Edmund Beecher Wilson denied the originality of
Guyer's suggestions in his paper of 1902 dealing with the
relationship
between the chromosome behavior and Mendel's traits (Wilson 1902). Stressing the importance of the conclusions reached by Cannon's
paper and Sutton's first article On the Morphology of the Chromosome
Group in Brachystola magna, that the separation of paternal and maternal elements in separate germ cells might explain the Mendelian
principle on a physical basis, Wilson quoted Guyer indicating that
his
suggestion of a split between parental chromosomes was not
explained in his 'hypothesis of irregularities in the
maturationdivisions'
(Wilson 1902, 992). In our opinion, Wilson did not
understand Guyer's earlier suggestion for some unknown reasons. It
seems obvious from the text of the Spermatogenesis that this author
stressed many times how his splitting hypothesis of the parental chromosomes in germ cells of hybrid from very different parents
might occur in the offsprings resulting from normal crosses.
Conclusion
It seems clear that Guyer provided, in the full ignorance of
Mendel's law, relevant ideas on the process of meiosis, which were
to
be taken into account later by Cannon and Sutton, two cytologists
who are historically quoted as having been the first to stress the
parallel between the behaviour of Mendel's particles and the
behaviour of the chromosomes. In assuming on a priori grounds that
the chromosome behaviour observed in the germ cells of hybrids from
very different forms would reflect the chromosome behaviour in
normal hybrids, Guyer suggested for the first time before
Montgomery's idea that the synapsis stage in the hybrids was the
result
of a conjugation of maternal and paternal chromosomes and that a segregation mechanism of the parental chromosomes might occur in
the final gametes. Interestingly, he perceived a clear parallel
between
the meiotic behaviour of pairs of chromosomes and the behaviour of the plumage colour trait in stressing in a Mendelian way the
'purity'
of the final gamete. Each gamete might contain one of the both
parental qualities, or in other words, all the maternal or all the
paternal chromosomes. Furthermore, in combining a statistical
approach from hybridisation results with cytological observations,
he
proposed a coherent chromosome behaviour mechanism involving
maternal and paternal chromosome separation in the reduction
division and their random recombination during fertilization to explain logically the offspring variation and the phenomenon of
reversion in the third generation.
In our opinion, this cytological chromosome mechanism is the same
as the one proposed by William Cannon in 1902, and exerted an
influence on Sutton's chromosome theory of 1903, which expressed for the first time the independent assortment of the chromosomes. To our
knowledge, Guyer's suggestion was also the first coherent attempt to
establish a parallel between chromosome behaviour in meiosis and hereditary character behaviour observed in successive generations.
It might be stressed how these cytological suggestions in Guyer's
dissertation were taken into account by the scientific community
through journals such as Science or the Bulletin of the University
of
Cincinnati. However, as the account in Science by 1900 was very
brief, Guyer's important suggestions were taken into consideration
by the scientific community later by 1902 when a whole summary
of his dissertation was published. Thus it seems clear that Guyer's
original suggestions in his thesis on pigeons were probably unknown because no account of his work had been published in
scientific journal. To our knowledge, nobody quoted this original
work.
In our opinion, this example of an early coherent cytological
suggestion attempting to explain offspring variation in crossing
supports the importance of the cytology in the late 19th century to
conceptualise a chromosome theory of heredity. The conception of the
central role of the chromatin as the physical basis of inheritance
and
the recognition of the individuality of the chromosomes as the
carriers
of hereditary traits led Guyer to support his interesting chromosome
mechanism hypothesis, although he did not know at the time about
the theoretical principles of Mendel's work. It is obvious that the
Mendelian principles undeniably provided some light to the
establishment of the final step in the chromosome theory of heredity by 1903 with the correct assumption of the universality of the
process
involving an independent segregation of the chromosomes in the germ
cells. However, we stress how this earlier and coherent suggestion
of
Guyer by 1900 supports Robert Olby's claim that cytologists would
have probably understood the laws of heredity by themselves in their
ongoing research to find a physical basis for heredity (Olby 1966,136).
Finally, the originality of Guyer's work relating hybridisation
results
with cytological observations illustrates the importance of the
connection between hybridization studies and cytology in United States at this time (Paul and Kimmelman 1988). This explains why it
was an American biologist, Sutton, who the first pointed out
correctly
the chromosome explanation of Mendel's laws in 1903. Boveri only published the same conclusions one year later in 1904.
Acknowledgements
We are grateful to Dr. Marc Ratcliff and M. Jacques Bungener for
their useful suggestions and advice on the manuscript. We would also
like to express our gratitude to two anonymous reviewers for
constructive comments on an earlier version of this manuscript.
References
Bateson W., Saunders E.R., 1902, Reports to the Evolution Committee:
Reports I,
London: Harrison & Sons.
Baxter A., Farley J., 1979, 'Mendel and Meiosis', Journal of the
History of Biology,
12: 137-173.
Boveri T., 1904, Ergebnisse Uber die Konstitution der Chromatischen
Substanz des
Zellkerns, Jena: Fisher.
Bulmer M., 1999, 'The development of Francis Galton's ideas on the
mechanism of
heredity', Journal of the History of Biology, 32: 263-292.
Cannon W.A., 1902, 'A Cytological Basis for the Mendelian Laws',
Bulletin of the
Torrey Botanical Club, 29: 657-661.
Cannon W.A., 1903, 'Studies in Plant Hybrids: The Spermatogenesis of
Hybrid
Cotton', Bulletin of the Torrey Botanical Club, 30: 133-172.
Cannon W.A., 1904, Some Cytological Aspects of Hybrids, (Proceedings
of
International Conference on Plant Breeding and Hybridization held in
the the
rooms of the American Institute of the City of New York and in the
Museum
building of the New York Botanical Garden, September 30 and October
1 and
2, 1902, 89-92).
Churchill F.B., 1970, 'Hertwig, Weismann, and the Meaning of
Reduction Division
circa 1890', Isis, 209: 429-457.
Coleman W., 1965, 'Cell, Nucleus, and Inheritance: An Historical
Study', Proceedings of the American Philosophical Society, 109: 124-158.
Corcos A.F., Monaghan F.V., 1997, 'Correns, an independent discover
of
Mendelism? II. Was Correns a real interpreter of Mendel's paper?',
The Journal
of Heredity, 78: 404-405.
Darden L., 1977, 'William Bateson and the Promise of Mendelism',
Journal of the
History of Biology, 10: 87-106.
Darden L., 1991, Theory Change in Science: Strategies from Mendelian
Genetics,
New York: Oxford University Press.
Davenport C.B., 1901, 'Mendel's Law of Dichotomy in Hybrids',
Biological Bulletin,
2: 307-310.
Gayon J., 1993, 'Quelque Temps Apres, Au Debut du Siecle', Les
Cahiers de Science
& Vie, 15: 36-44.
Guyer M.F., 1899, 'Ovarian structure in an abnormal pigeon',
Zoological Bulletin, 2:
211-224.
Guyer M.F., 1900a, Spermatogenesis of Normal and of Hybrid Pigeons,
Chicago:
University of Chicago.
Guyer M.F., 1900b, 'Spermatogenesis in Hybrid pigeons', Science, 11:
248-249.
Guyer M.F., 1902, Hybridism and the Germ-Cell, Cincinnati:
Cincinnati University
Press (Bulletin of the University of Cincinnati 21, series 2, vol.
2).
Guyer M.F., 1903a, 'The Germ Cell and the Results of Mendel', The
Cincinnati
Lancet-Clinic, 53: 490-491.
Guyer M.F., 1903b, Spermatogenesis of Normal and of Hybrid Pigeons,
Cincinnati:
Cincinnati University Press (Bulletin of the University of
Cincinnati 22, series 2,
vol. 3).
Guyer M.F., 1907, 'Do Offspring Inherit Equally From Each Parent?',
Science, 25:
1006-1010.
Guyer M.F., 1911, 'Nucleus and Cytoplasm in Heredity', The American
Naturalist,
45: 284-305.
Maienschein J., 1987, 'Heredity/Development in the United States,
circa 1900', History and Philosophy of the Life Sciences, 9: 79-93.
Mayr E., 1976, 'Charles Otis Whitman'. In: Gillispie C.C. (ed.),
Dictionary of
Scientific Biography, vol. 14, New York: Charles Scribner's Sons,
313-315.
McClung C.E., 1902, 'The Accessory Chromosome - Sex Determinant?',
Biological
Bulletin, 3: 43-84.
McKusick V.A., 1960, 'Walter S. Sutton and the physical basis of
Mendelism', Bulletin of the History of Medicine, 34: 487-497.
Montgomery T.H., 1901, 'A Study of the Chromosomes of the Germ Cells
of
Metazoa', Transactions of the American Philosophical Society, 20:
154-236.
Montgomery T.H., 1903, 'Some Observations and Considerations upon
the
Maturation Phenomena of the Germ Cells', Biological Bulletin, 6:
137-163.
Montgomery T.H., 1904, 'The Main Facts in Regard to the Cellular
Basis of
Heredity', Proceedings of the American Philosophical Society, 43:
5-14.
Murehead Macfarlane J., 1891, 'A Comparison of the Minute Structure
of Plant
Hybrids with that of their Parents, and its Bearing on Biological
Problems', Transactions of the Royal Society of Edinburgh, 37: 203-283.
Noland L.E., 1972, 'Michael Frederic Guyer'. In : Gillispie C.C.
(ed.), Dictionary of
Scientific Biography, vol. 5, New York: Charles Scribner's Sons,
598-599.
Olby R., 1966, The Origins of Mendelism, London: Constable.
Paul D.B., Kimmelman B.A., 1988, 'Mendel in America: Theory and
Practice, 1900-1919'. In : Rainger R., Benson K.R., Maienschein J. (eds),
The
American
Development of Biology, University of Pennsylvania Press, 281-310.
Richmond M.L., 2001, 'British cell theory on the eve of genetics',
Endeavour, 25: 55-60.
Riddle O. (ed), 1919, Orthogenic Evolution in Pigeons. Posthumous
Works of C. O.
Whitman, 2 vols, Washington: The Carnegie Institution of Washington.
Sapp J., 1987, Beyond the Gene: Cytoplasmic Inheritance and the
Struggle for
Authority in Genetics, New York, Oxford: Oxford University Press.
Shor E.N., 1971, 'Charles Benedict Davenport'. In : Gillispie C.C.
(ed.), Dictionary
of Scientific Biography, vol. 3, New York: Charles Scribner's Sons,
589-591.
Sutton W., 1902, 'On the Morphology of the Chromosome Group in
Brachystola
Magna', Biological Bulletin, 4: 24-39.
Sutton W., 1903, 'The Chromosomes in Heredity', Biological Bulletin,
4: 231-251.
Voeller B.R. (ed.), 1968, The Chromosome Theory of Inheritance:
Classic Papers in
Development and Heredity, New York: Appleton-Century-Crofts.
de Vries H., 1900, 'Sur la loi de disjonction hybrides', Comptes
Rendus de
l'AcadEmie des Sciences, 130: 845-847.
Wilson E.B., 1900, The Cell in Development and Inheritance, London:
Macmillan &
Co.
Wilson E.B., 1902, 'Mendel's Principles of Heredity and the
Maturation of the
Germ Cells', Science, 16: 991-993.
![]()
This web-page was created August 2011 by D. R. Forsdyke and was last edited 10 Nov 2020