|
Two Levels of Information in DNA Relationship of Romanes' "Intrinsic" Variability of the Reproductive System, and Bateson's "Residue," to the Species-Dependent Component of the Base Composition, (C+G)%
D. R. Forsdyke (1999) J.
Theoretical Biology 201, 47-61. [Footnote: Unless otherwise stated, italics and words within square brackets in quotations, are those of the present author, and should not be attributed to the person quoted.]
|
![]()
![]()
In 1886 Charles Darwin's research associate George Romanes published a paper entitled "Physiological Selection: An Additional Suggestion on the Origin of Species". This was criticized by his Victorian contemporaries and largely ignored by those who followed. However, the recent recognition of two levels of information in DNA suggests that Romanes had solved the major problems with Darwin's theory. It was apparent from the outset that the form of reproductive isolation likely to apply most generally to initial species divergence (hybrid sterility), would depend on differences, not in "primary" information ("genic"), but in "secondary" information ("chromosomal"). This viewpoint, further elaborated by Bateson & Saunders (1902), White (1978), and King (1993), is criticised by the genic school (Coyne & Orr, 1998) because it requires visible differences between chromosomes, and appears not to explain Haldane's rule. However, chromosomal differentiation with respect to the species-dependent component of base composition [(C+G)%; Forsdyke, 1996] appears to resolve these problems. Because it explained so much, it was easy to believe that the genic viewpoint explained everything. Romanes and Bateson thought otherwise. We are only just beginning to recognize what they were trying to tell us. |
| In 1978 Micheal White (Click Here), a major advocate of chromosomal speciation, concluded his Modes of Speciation by observing: |
|
|
| M.J.D. White 1910-1983 |
Thus White predicted that by now the modern era of speciation research, which began in 1859 with Charles Darwin's The Origin of Species, should be complete. I here propose that White may have been quite close to the mark. I show that a modification of his "chromosomal" viewpoint (Forsdyke, 1996, 1998) meets the criticisms raised by those who favour the alternative "genic" viewpoint (Coyne & Orr, 1998). I also propose that we now have a "new synthesis" not only of evolutionary theory, but also of its history (Forsdyke, 1999a). Just as the full understanding of a subject requires an understanding of its history, so the full understanding of its history requires an understanding of the subject. John Lesch observed in 1975 that: |
"The development of evolutionary theory in the two decades from Darwin's death to the turn of the century remains very largely terra incognita for the historian." |
| William Provine (1986) concluded similarly that: |
| "Evolutionary biology in the period 1859-1925 is extraordinary complex". |
I suggest here that successive studies in this period, first of Darwin's research associate, George Romanes (1886, 1887, 1897). and then of the biologist William Bateson (Bateson & Saunders, 1902; Bateson, Saunders & Punnett, 1904; Bateson, 1904, 1909a,b, 1913, 1922, 1924), can now be understood after a century of neglect. |
The idea of barriers against members of other species is apparent in the context of infectious disease. The barriers are both external (e.g. hygienic measures), and internal (e.g. the immune response). Internally, our bodies ("self") can detect and destroy members of other species ("not-self" viruses, bacteria, protozoa). Another, no less subtle form of self/not-self discrimination, involves our detection of a mate ("near-self") who will be our "physiological complement" such that the union will produce healthy children ("hybrids"). An incestuous relationship with a close relative ("too near-self") will probably result in less healthy children. On the other hand, extreme out-breeding, such as with an ape (not-self), is prohibited by species barriers. There is more to this than just the inability to copulate (the gamete transfer barrier). There are both external components (e.g. geographical isolation, mate choice), and internal components. Even if the male sperm could meet and fuse with the female ovum, the resulting cell (zygote) might be unable to grow into an adult organism (developmental barrier resulting in hybrid inviability). Even if these transfer and developmental barriers were overcome, in the gonad the two sets of parental chromosomes might be unable to pair for the production of gametes (gonadal barrier resulting in hybrid sterility). Speciation is the process by which such barriers are established. The issues are complex: |
|
Semantic problems cloud these issues, so I have tried to use plain language and a limited number of defined terms (see Box. 1). These definitions may be controversial, but it is to be recalled that progress in science (e. g. physics at the turn of the 19th century) sometimes requires that conventional definitions be challenged. |
| Evolutionary event.
Evolution results from the accumulation of many evolutionary "events", each involving some or all of four processes: variation, heredity (inheritance), phenotypic selection, reproductive isolation. An evolutionary event begins with variation and is followed by the process of heredity continuing through the generations. Selection may follow variation. |
| Selection.
The words "selection" and "isolation" have led to confusion. The words have the same meaning, although one may be preferred in a particular context. Selection is something which can be done when there is more than one of something. If there is only one of something, it is already selected (or isolated). In much of the literature, "selection" when used alone often means phenotypic selection; "isolation" when used alone often means reproductive selection. |
| Phenotypic selection.
There is positive selection of members of the species with advantageous variations, and negative selection of members of the species with disadvantageous variations. This isolation on the basis of phenotypic adaptations (positive or negative) is the type of selection with which Darwin was primarily concerned. In this respect he and others spoke of "survival of the fittest" and the "struggle for existence". He used the term "natural selection", to distinguish this form of phenotypic selection from the artificial selection carried out by the domestic breeder. However, reproductive isolation/selection can also be natural or artificial. |
| Reproductive isolation.
Variation, heredity and phenotypic selection can operate so that advantageous phenotypic characteristics would tend to increase in a population, and disadvantageous phenotypic characteristics would tend to decrease in a population. They lead to the adaptation of species, and operate primarily at the level of the individual member of the species (individual selection). However, for the origin of species (i. e. a divergence between two population groups), a fourth process, "reproductive isolation", is needed. To originate species, groups have to be selected. Speciation is a form of group selection. A group is a set of units. For our purposes, if there is only one unit in a set it is an individual, and does not constitute a group. A group whose members reproduce only asexually are, by definition, reproductively isolated from other groups. |
| Species.
The most secure definition of a biological species is a group of organisms which is reproductively isolated from other groups of organisms. Members of the group are infertile (sterile) with all other organisms, except those organisms which belong to the group. Degrees of infertility are characteristic of organisms at the species interface (i.e. potential incipient species). |
| Hybrid.
The offspring of crosses between members of two anatomically distinct lines (varieties, races) are often referred to as hybrids. Since the definition of species is based primarily on reproductive performance, rather than anatomical difference, this can lead to confusion. For present purposes any offspring is the hybrid of its parents if the parents differ genomically. If the offspring is more fit than the parents there is hybrid vigour (positive heterosis). If the offspring were less fit, this negative heterosis could include hybrid inviability or sterility. The only possible human offspring not a "hybrid" would a female who inherited absolutely identical sequences from both parents. Human males are "hybrids" since, even if the autosomes were identical, the sex chromosomes would differ. |
3. Problems with Darwin's theory
 |
Those close to Darwin were quick to point to problems with his theory. Joseph Hooker (1862) agreed that heredity (like produces like) was fundamental to the theory, but maintained that variation (like does not produce like) was equally fundamental; furthermore, variation was an autonomous force, analogous to gravity, largely independent of environmental factors. |
While Darwin's "natural selection" required what was later called "survival of the fittest", Hooker and others proposed what Romanes was to call "survival of the fit" (survival of the survivors, or survival of those not manifestly unfit), by a process independent of natural selection (Hooker, 1962; Gulick, 1872; Delboeuf, 1877). This is now best equated with "random drift". |
|
Thomas Huxley (1863) thought that Darwin had not adequately explained hybrid sterility. Thus, the cross of a horse with an ass is usually fertile ("first crosses" succeed), but the hybrid progeny, a mule, is usually sterile ("second crosses" fail). Gamete production in the maldeveloped gonads of the mule is impaired. |
 |
In 1862 Darwin made notes on hybrid sterility, which have recently been published; these show him toying with what Romanes was later to call "physiological selection"; however, Darwin soon gave up declaring "will not do." |
|
.
Romanes focussed on the sterility barrier, which often appears to be the only barrier separating members of closely related (allied) species, and, indeed, seems the most useful criterion for distinguishing such species (Box 1). He suggested that, like cells of all other organs and tissues, germ line cells might undergo random variations. Variations, for example, in height or eye colour were familiar to everyone. In 1886 no one knew what caused variations, but no one doubted their existence. Romanes emphasized one possible class of variation affecting germ line cells, which would make an organism less fertile (more sterile) with other members of the species but would not directly influence somatic characters. Normally the loss of fertility would be highly disadvantageous because the organism would leave no offspring. However, he argued that if two organisms underwent the same type of variation, they would still be fertile between themselves. They would be "physiological complements" (Wallace, 1886; Romanes, 1887, 1897). Hence, at any time-point a species would consist of the parental group (comprising the majority of species members), and numerous small variant groups. Members of each variant group would be less fertile with members of the parental group and of the other variant groups. Members of each group, to varying degrees, would be reproductively isolated from members of the other groups, but remain quite fertile with each other. Full hybrid sterility would be an extreme manifestation of the phenomenon. |
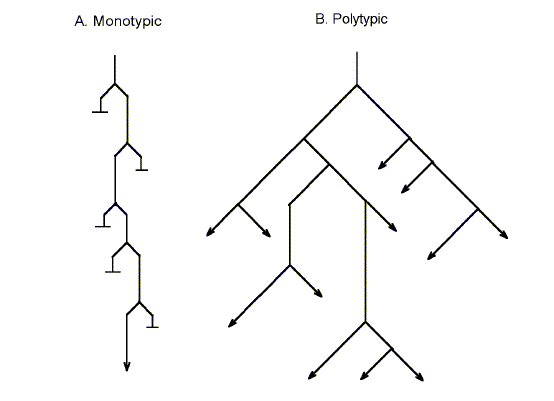
Romanes argued that irrespective of any selective factors in the environment (no survival of the fittest required), members of the reproductively variant groups would tend also to be somatically variant, just as members of the parental group would also tend to be somatically variant (e.g. changes in height or eye colour). However, in the absence of any environmental selection, somatic variations of members of the large parental group would not be sustained because these members were crossing freely with each other, a process which would tend to blend and neutralize variations. For example, although humans vary in height (a multigenic character), tall people cross with small people and the average height tends to remain constant. On the other hand, variations within a small variant group would not be subjected to this "swamping" effect due to blending with members of the parental group (i.e. there would be less non-variants to dilute the variation). Because, and only because, of this reproductive isolation, the variation would be sustained. If, by chance, the variation happened to confer some advantage on members of the variant group, relative to members of the parental group, this would be a bonus. The selective advantage conferred would allow the commencement of classical Darwinian natural selection. The major difference between the old Darwinian formulation (natural selection), and the new Romanesian formulation ("physiological selection") was that in the first case, natural selection preceded reproductive isolation, where in the second case reproductive isolation preceded natural selection. Reproductive isolation, in any shape or form, would suffice, but Romanes held that the most usual form of initial isolation would result from physiological selection; this would first be manifest as partial, and then proceed to complete, hybrid sterility. Of course, in a small population it would be very unlikely that a male and female whose gonads had undergone the same rare variation would appear at the same time. Furthermore, although in a large population two individuals with the same rare gonadal variation might appear, they would be unlikely to encounter each other. Thus, neither in small nor in large populations would there be much chance of the variation being perpetuated. Indeed, as Romanes pointed out, successful speciation seemed rare. This type of evolution was referred to as "polytypic" in that it allowed a branching into divergent lines, a process to be contrasted with the linear "monotypic" evolution brought about by natural selection acting alone on sexual populations. Romanes pointed out that in monotypic evolution reproductive isolation was still a factor, being achieved by the most secure method possible, the death of the non-adapted (Fig. 1). |
|
|
|
FIG. 1. The distinction between (A) unbranching "monotypic" evolution, and (B) branching "polytypic" evolution. |
6. "Intrinsic" cause of hybrid sterility
In 1886 Romanes ascribed physiological selection to a spontaneously arising "peculiarity" of the reproductive system and regarded his studies as merely paving the way for future investigations: |
"My suggested explanation of the origin of species opens up another and more ultimate problem, namely, granted that species have originated in the way supposed, what have been the causes of the particular kind of variation in the reproductive system which the theory requires?" |
To give his Victorian audience a concrete example of one form of physiological selection, Romanes mentioned a change in the time of flowering of a plant, which would restrict the plant to fertilization only by other members of the species which had undergone the same change in flowering time: |
"Suppose the variation in the reproductive system is such that the season of flowering, or of pairing, becomes either advanced or retarded. ...Some individuals living on the same geographical area as the rest of their species, have varied in their reproductive system, so that they can only propagate with each other. They are thus perfectly fertile inter se, while absolutely sterile with all the other members of their species. This particular variation being communicated by inheritance to their progeny, there would soon arise on the same area, ... two varieties of the same species, each perfectly fertile within its own limits, while absolutely sterile with one another. That is to say, there has arisen between these two varieties a barrier of intercrossing which is quite as effectual as a thousand miles of ocean; the only difference is that the barrier, instead of being geographical, is physiological." |
Today we would regard such cases as most likely to result from primary variations (mutations) in specific genes which affect the time of flowering, thus creating a gamete transfer barrier; this would be a "genic" means of achieving reproductive isolation. However, Romanes (1886) proposed that something more than this "extrinsic" cause was involved in the general case of formation of new species by physiological selection. |
"In many cases, no doubt, this particular ... variation, has been caused by the season of flowering or of pairing having been either advanced or retarded in a section of a species, or to sundry other influences of an extrinsic kind; but probably in a still greater number of cases it has been due to what I have called intrinsic causes, or to the "spontaneous" variability of the reproductive system itself." |
The change in time of flowering ("extrinsic" cause) was not associated with degeneration of the gonad, unlike hybrid sterility ("intrinsic" cause). Thus, pollen from an early-flowering variant, if saved, was often found successfully to fertilize a late-flowering variant. Similarly, variants in pairing time (cicadas, moths) can also be successfully crossed (Bateson, 1913). |
7. Chromosomal basis of hybrid sterility
Coinciding with the reemergence of Mendel's work at the turn of the century, microscopical studies of chromosomes in the gonads of sterile hybrids by Michael Guyer (1902) and others revealed the cytological basis of hybrid sterility. Normally, the set of chromosomes carrying genes contributed by the mother pair, on a one-to-one basis, with the complementary set of chromosomes carrying the genes contributed by the father. Close identity is essential for this pairing, which is required so that subsequent cell division and multiplication can proceed, resulting in the formation of gametes. In hybrid sterility the pairing is defective and this is associated with a degenerate gonad. |
|
 |
Thus the question of the origin of hybrid sterility, Romanes' "intrinsic" cause of variation leading to the origin of species, progressed to the question of the nature of the chromosomal incompatibilities which prevented pairing, or conversely, the nature of the chromosomal compatibilities which permitted pairing. Reminiscent of Romanes' position, the English biologist William Bateson (1913) interpreted hybrid sterility as reflecting complementary chromosomal "factors": |
"Though we cannot strictly define species, they yet have properties which varieties have not, and ... the distinction is not merely a matter of degree. The first step is to discover the nature of the factors which by their complementary action inhibit the critical divisions and so cause the sterility of the hybrid." |
Romanes and Bateson used the word "complementary" in converse senses. Romanes' "physiological complements" were organisms with reproductive systems which were compatible, so that offspring would be produced. Bateson (1913) held that organisms were incompatible and failed to complement in this way, either because the reproductive system of , |
|
"each is lacking in one of two complementary elements, or that each possesses a factor with an inhibitory effect". |
| Hybrid sterility was to be thought of |
|
"as something apart, and a manifestation which distinguishes certain kinds of unions in a very special way." |
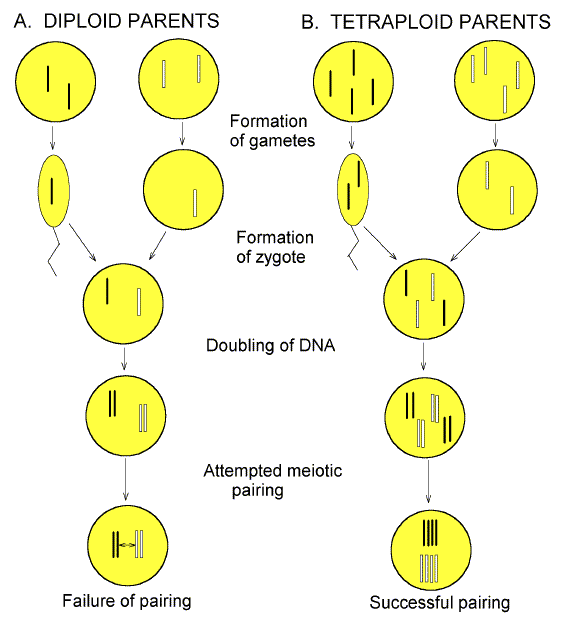
That hybrid sterility might result from a failure of the pairing of complementary chromosomes was later brought home forcefully by experiments showing that hybrid sterility could be "cured" in certain organisms by artificially doubling the number of chromosomes (Winge, 1917; Darlington, 1932). This was because each chromosome now had a partner with which it could pair (i.e. chromosomes from the mother could pair with each other, and chromosomes from the father could pair with each other). If the sterility had been due to a defect in particular genes, this "cure" would be unlikely (Figure 2). |
|
|
FIG. 2. Polyploidy "cures" hybrid sterility. Chromosomes from the father (black), and from the mother (white), are brought together as gametes to form zygotes (no transfer barrier). Development is unimpaired (no hybrid inviability). However, in the case of diploid parents (A), pairing at meiosis is impaired due to differences between the parental chromosomes, so that gamete formation fails (hybrid sterility). In the case of tetraploid parents (B), gametes are diploid, so that pairing at meiosis can occur. |
8. Two levels of genetic information
Bateson was much occupied with overcoming the opposition to Mendel (Olby, 1985), and does not appear to have been influenced by Romanes' work. He seems not to have fully recogized the importance of hybrid sterility until some time after 1904 (Forsdyke 1999a). However, as early as 1902 he and Saunders were citing a 1900 abstract by Guyer and indicating a sense of the need for two levels of genetic information. Bateson later (1909a) cited both Guyer's 1900 thesis and his 1902 paper. The two levels of genetic information eventually became known as "genic" and "chromosomal", and seem to correspond, respectively, with Romanes' "extrinsic" and "intrinsic" causes of the variation leading to the generation of "physiological complements". In a Report to the Evolution Committee of the Royal Society, Bateson and Saunders noted (1902): |
"Though some degree of sterility on crossing is only one of the diverse properties which may be associated with Specific [species] difference, the relation of such sterility to Mendelian phenomena must be a subject of careful enquiry. ...The sterility of certain crosses is merely an indication that they cannot divide up the [parental] characters among their gametes. If the parental characters, however dissimilar, can be split up [segregated], the gametes can be formed, and the inability to form gametes may mean that the process of resolution [separation of characters] cannot be carried out. ... That the sterility of hybrids is generally connected in some way with inability to form germ cells correctly, ... is fairly clear." (Bateson's italics) |
That Guyer's observed chromosomal changes might be the cause of, rather than an effect of, hybrid sterility, was noted by Walter Sutton in 1903, but Bateson and Saunders went further: |
"Has a given organism a fixed number of unit-characters? Can we rightly conceive of the whole organism as composed of such unit characters, or is there some residue -- a basis -- upon which the unit characters are imposed? We know, of course, that we cannot isolate this residue from the unit characters. We cannot conceive of a pea, for example, that has no height, no colour, and so on; if all these were removed there would be no living organism left. But while we know that all these characters can be interchanged [between members of a species], we are bound to ask is there something not thus interchangeable? And if so, what is it? We are thus brought to face the further question of the bearing of Mendelian facts on the nature of species. The conception of Species, however we may formulate it, can hardly be supposed to attach to allelomorphic ... varieties [conventional genes]. We may be driven to conceive "Species" as a phenomenon belonging to that "residue" spoken of above, but on the other hand we get a clearer conception of the nature of sterility on crossing [hybrid sterility]." |
There is here a distinction between "unit characters" (conventional "genic" characters), and another form of genetic information, a "residue" or "basis". Unlike the unit characters which can be "interchanged" over the generations (e.g. smallness to tallness in peas), the "residue" is held to be unchangeably linked to the species. A distinction between "sporadic" (genic) forms, and what we would now call "chromosomal" forms of hybrid sterility was later drawn by Bateson, Saunders and Punnett in 1904 when commenting on sporadic mutants with withered anthers: |
"[Withered] anthers were seen from time to time in many families, though commonly confined to individual flowers. This sporadic sterility has not been particularly studied. It is of interest to compare this example of the definite appearance of sterility ... with the familiar [regular] occurrence of sterility in cross- breds [hybrids]. Such a phenomenon has often been supposed to indicate remoteness of kinship, yet here a closely comparable effect occurs ... as the result of a cross between two types which must be nearly related. Mr. Gregory in a careful examination of the pollen-genesis [in the case of crosses between nearly related types] found that the divisions were normal up to the reduction division, when the chromosomes formed shapeless knots and entanglements, failing to divide [i.e. the disruption of meiosis was similar to that seen in crosses between types remote in kinship]." |
In 1913 Bateson dismissed sterility associated with such sporadic anther variants in plants as being due to a "simple recessive character," again implying, as had Romanes, that sterility due to defects in one or a few defined characters ("genic") was unlikely to be of general significance for hybrid sterility as affecting the evolution of species. Under the heading "Possible limits to recombination" [barriers to sex] Bateson (1909a) noted: |
"It will probably occur to many that there are limits to these possibilities of transference [of genic characters], and so undoubtedly there are. The detection of these limits [which include barriers affecting speciation] is one of the more important tasks still awaiting us. ... We must surely expect that these transferable characters [conventional genic characters] are attached to, implanted upon, some basal organization, and the attributes or powers which collectively form that residue may perhaps be distinguishable from the transferable qualities. The detection of the limits thus set [by the basal organization or residue] upon the inter-changeability of characters [from parents to offspring over the generations] would be a discovery of high importance and would have a most direct bearing on the problem of the ultimate nature of Species." |
Bateson remained convinced of the importance of two fundamental levels of genetic information to the end of his life. In 1922 he declared that he was: |
"...Left with the conviction that some part of the chain of reasoning is missing. ... [Genetic] analysis has revealed hosts of transferable characters [genes]. Their combinations suffice to supply an abundance of [anatomically differentiated pheno]types which might pass for new species, and certainly would be so classed if they were met with in nature. Yet critically tested [by cross-breeding], we find that they are not distinct species and we have not reason to suppose that any accumulation of characters of the same order [transferable characters amenable to conventional genetic analysis] would cumulate in the production of distinct species. ... Specific difference, [that which is responsible for speciation] therefore must be regarded as probably attached to the base upon which these transferables are implanted, of which we know absolutely nothing at all." |
| In 1924 he returned to the same theme: |
"We have determined the transferable [genic] characters as one group, and we no longer confound them with the essential elements [another group] conferring specificity [speciation]. Segregation [of genic characters] is of course often seen in species crosses, but as to the behaviour of these critical elements [those conferring species specificity] we know as yet very little." |
Perhaps it was because Bateson was seeking so much more from chromosomes than the mere transfer of genic characters that to many he appeared "conservative" and "confused" regarding the role of chromosomes (Coleman, 1970; Orr, 1996). As Bateson's words are read, the modern reader can have in mind the DNA molecule (discussed below). His contemporaries did not have this advantage. |
Romanes' closest supporter had been the American missionary John Gulick. The latter's son, Addison, continued the crusade. In his 1932 biography of his father he wrote: |
"The problem of physiological isolation has received but little attention during the last two decades, but the modern picture of the mechanism of reproduction and the physico-chemical processes that it involves would have a very large influence on any consideration of the subject today. The immediate cause of infertility between species would be sought at the present time in either chromosomal incompatibilities or maladjustments of a serological nature .... From several viewpoints the old proposition of Romanes seems today exceedingly plausible, that a rather trivial mutation or group of mutations might set up a barrier of sterility within a species, and cause the two portions to diverge thereafter into well-marked new species". |
In the same year Cyril Darlington, in his Recent Advances in Cytology came to recognize that some form of reproductive isolation was likely to precede natural selection:, |
"It is clear that discontinuity between species is due to isolation being followed by independent variation. Isolation is doubtless often brought about by [geographical] distribution, by change of habit, or by genetic sterility factors; it is also brought about by structural and numerical changes in the chromosomes." |
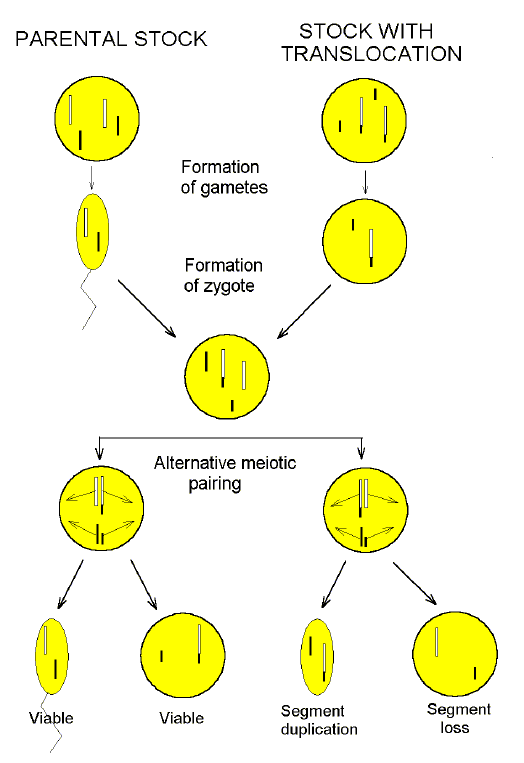
There is here a spelling out of the important distinction between isolation due to the action of specific genes ("genic" sterility), and due to changes in chromosomes ("chromosomal" sterility). In the latter case Darlington meant impaired meiotic chromosome pairing due to microscopically visible changes which we will call "macromutations" (e.g. translocations, inversions, deletions, changes in chromosome numbers) in one of the pairing partners. If the pairing partners had the same macromutation, then pairing would not be impaired and the hybrid would be fertile. A version of this, advanced by Addison Gulick (1932), is shown in Figure 3. |
|
|
FIG. 3. Addison Gulick's proposed basis of "chromosomal" sterility due to a translocation ("macromutation"). There has been a translocation of a small piece of one chromosome (black) to another chromosome (white). There is no transfer barrier or hybrid inviability. When pairing is attempted at meiosis, chromosomal segments assort randomly. Half the resulting gametes will have the full chromosomal complement, but the other half will have lost or gained large segments. Various "check-point" mechanisms may prevent these gametes forming; even if formed, any resulting zygotes should be inviable. Gulick pointed out that if there is one translocation, only half the gametes will be viable; for two translocations, only a quarter would be viable; for N translocations, only 1/(1+N) gametes would be viable. [The term "macromutation" should be understood in context, since it is used in a variety of senses in the genetics literature.] |
The chromosomal viewpoint found a strong advocate in White. In his Modes of Speciation in 1978 he concluded that: |
"Over 90% (and probably over 98%) of all speciation events are accompanied by karyotic changes [chromosomal macromutations], and ... in the majority of cases the structural chromosomal rearrangements have played a primary role in initiating divergence". |
Shortly thereafter, evolutionist Stephen Gould (1980) in a paper entitled "Is a new and general theory of evolution emerging?" noted a new readiness to entertain "chromosomal alterations as isolating mechanisms", and pointed out that: |
"Some of the new models ... regard reproductive isolation as potentially primary and non-adaptive rather than secondary and adaptive ... . In ... chromosomal speciation, reproductive isolation comes first and cannot be considered an adaptation at all. It is a stochastic event that establishes a species by the technical definition of reproductive isolation. To be sure, the later success of this species in competition may depend on its subsequent acquisition of adaptations; but the origin itself may be non-adaptive. We can, in fact, reverse the conventional view and argue that speciation, by forming new entities stochastically, provides raw material for selection". |
Another strong advocate of the chromosomal viewpoint was Max King. However, in his 1993 book Species Evolution: The Role of Chromosome Change he acknowledged various criticisms of the viewpoint. These have been listed by Jerry Coyne and Allen Orr (1998), who strongly favour the genic viewpoint (Box 2). Some of these criticisms are valid if chromosomal speciation is held strictly to require large chromosomal changes ("macromutations"). I will now outline the "new synthesis", described fully elsewhere (Forsdyke, 1996, 1998) and show how it meets these criticisms. |
1. "Many species producing sterile hybrids ... do not differ in chromosomal arrangement. ... Many of the taxa showing fixed chromosomal differences are old, and may have accumulated many -- if not all -- of their chromosomal differences after the actual speciation event." |
2. "Chromosomal speciation...depends critically on the semisterility of hybrids who are heterozygous for chromosome rearrangements. It is not widely appreciated, however, that the heterozygous rearrangements theoretically expected to be deleterious (e.g. fusions and pericentric inversions) in reality often enjoy normal fitness." |
3. "Even in species hybrids whose chromosomes fail to pair during meiosis, we do not know whether this failure is caused by differences in chromosome rearrangement of differences in genes." |
4. "Direct genetic analyses over the last decade have shown conclusively that postzygotic isolation in animals is typically caused by genes, not by large chromosomal rearrangements. Many of these genes have been genetically characterized or even cloned." |
5. "Although sterile hybrids often suffer meiotic pairing problems, this sterility is often limited to the heterogametic sex [Haldane's rule]. This is not expected if sterility results from the disruption of chromosome pairing, which in most cases should afflict both males and females." |
10. Metaphors for an exclusionary role of secondary information
One form of information is often accompanied by another form which plays an exclusionary role protecting the first form. Thus, a radio signal containing a message ("primary information") is transmitted at a particular wavelength ("secondary information"). The signal contains both forms of information. The wavelength permits a receiver to discriminate between the accompanying primary message and other primary messages which would be transmitted at other wavelengths. Whereas the message information tends to be unique for each particular part of the signal, the wavelength information tends to be uniformly dispersed over every part of the signal. |
 |
In similar fashion, our spoken words (primary information) are accompanied by a uniformly dispersed accent (secondary information). Disparities in accent sometimes influence reproductive choices. In Shaw's 1913 play Pygmalion, the accent of the cockney flower-seller Eliza Doolittle makes it unlikely that she will marrying the "toff" Freddy Eynsford Hill, a man of some social standing (Forsdyke 1999a). |
We may regard Eliza's dropped Hs as "micromutations". Her secondary information, because of such micromutations, constitutes a reproductive barrier. Cockneys tend to marry cockneys and perpetuate cockney secondary information. The middle class tend to marry into the middle class and perpetuate middle class secondary information. Shaw tells us that Professor Higgins performed the appropriate experiment. He demonstrated that if the linguistic barrier were removed, then the reproductive barrier would also be removed. Eliza and Freddy lived happily-ever-after. We can say that they then possessed "linguistic complements", by which we mean that they approached linguistic identity. Alternatively, we can say that until Professor Higgins had performed his magic, their reciprocal differences, or "complementary factors" (Eliza's dropped Hs, and Freddy's included Hs) kept them reproductively isolated. We should also note that once the marriage knot had been tied (i.e. the barrier to reproduction formally removed), a good chance existed that, even if her accent subsequently regressed (diverged from Freddy's, so that linguistic complementarity was lost), the marriage (reproduction) would continue. Thus the initial secondary information (a reproductive barrier) needed only to be removed temporarily until a more substantial (legal) way of removing the barrier had been substituted. |
11. Two levels of information in DNA
Our genetic material, DNA, also contains two levels of information. The sequence of the four bases (A, C, G, T) provides primary (including "genic") information. However the relative proportions of the W bases (A and T) to the S bases (C and G) provides another level of information ("chromosomal"), constituting the uniform "accent" of DNA. Erwin Chargaff in 1951 showed that this base composition-dependent parameter, referred to as (C+G)%, is a species characteristic. A link with reproductive isolation was noted for strains of Tetrahymena by Noboru Sueoka (1961): |
"DNA base composition is a reflection of phylogenetic relationship. Furthermore, it is evident that those strains which mate with one another (i.e. strains within the same 'variety') have similar base compositions. Thus strains of variety 1 ..., which are freely intercrossed have similar mean GC content. ... If one compares the distribution of DNA molecules of Tetrahymena strains of different mean GC contents, it is clear that the difference in mean values is due to a rather uniform difference of GC content in individual molecules. In other words, assuming that strains of Tetrahymena have a common phylogenetic origin, when the GC content of DNA of a particular strain changes, all the molecules undergo increases or decreases of GC pairs in similar amounts. This result is consistent with the idea that the base composition is rather uniform not only among DNA molecules of an organism, but also with respect to different parts of a given molecule." |
A mechanism by which differences in GC [(C+G)%] might affect reproductive isolation arose from studies of Jih-Hsiang Chen and others (Chen et al., 1990; Forsdyke, 1998). They showed that extremely small fluctionations in (C+G)% would have a major influence on the ability of duplex DNA to extrude single-strand stem-loop structures. There is growing evidence that "kissing" interactions between the loops in such stem-loop structures play a fundamental role in the homology search which precedes the pairing of homologous maternal and paternal chromosomes at meiosis (Hunter et al., 1996; Rocco & Nicolas, 1996; Kleckner, 1997; Wong et al., 1998; Forsdyke, 1996, 1998). Thus, a small fluctuation in (C+G)% should profoundly affect the pairing of homologous chromosomes. This should activate various "checkpoint" mechanisms (Page and Orr-Weaver, 1996) resulting in a disruption of meiosis, impaired gametogenesis, and partial, and eventually complete, hybrid sterility. |
|
|
|
However, whatever the initial isolation process which led to the divergence between ancestral lines leading to, say, the modern elephant and mouse, it is clear that there are now more substantial barriers to reproduction between the two (e.g. a gamete transfer barrier). It would probably be fruitless to examine these species for molecular clues as to the way the initial divergence took place. In certain virus species, however, signs of the initial isolation process might still exist. They might have been unable to progress beyond the first barrier. This, indeed, may be the case with two virus species whose members have the potential to coexist in the same host cell (Yin & Hu, 1997); here they would exert a selection pressure on each other to sustain and expand any differences which might prevent recombination (otherwise they would recombine and destroy each other as individual species). As related elsewhere (Forsdyke, 1996), circumstantial evidence that differences in (C+G)% might exert anti-recombination effects first emerged from studies of insect viruses by Gerard Wyatt (1952), and there is now supporting data from analyses of herpes viruses (Schachtel et al., 1991), and of retroviruses (Bronson & Anderson, 1994). Other evidence, albeit also circumstantial, arose from studies of the uniting of genomes within a common nucleus to form novel polyploid species. Parental genomes of different (C+G)% can coexist for at least 6 million years yet remain recombinationally isolated. Matassi and coworkers (1991) propose this is a function of "nuclear architecture", but in the light of the present work it seems possible that the differences in (C+G)% would themselves be responsible for the isolation. Thus the data are consistent with DNA having at least two levels of information. The primary information includes localized "sentences", which are the genes encoding the various characteristics of an organism. Changes in these genes are responsible for conventional differences in phenotype. Changes in the (C+G)% secondary information, part of the "residue" upon which the genic "unit characters are imposed" (Bateson & Saunders, 1902), would provide an "intrinsic" molecular basis for Romanes' "physiological peculiarity" of the reproductive system leading to hybrid-sterility. The secondary information should match between two individuals ("physiological complements") in order for them to reproduce successfully (Sueoka 1961). A divergence in this secondary information, through accumulation of "micromutations" (single base changes), would initiate the speciation process (Forsdyke, 1996, 1998). We should note that Bateson's careful use of the general term "factors" (not "genes") allowed him to encompass any form of information (primary or secondary) which was contained in the chromosomes (Haldane, 1957). |
The first and second criticisms of Coyne and Orr (1998; Box 2) are that (i) species pairs, whose members when crossed produce sterile hybrids, often do not show chromosomal macromutational differences, and conversely (ii) when chromosomal macromutational differences are observed there may be no hybrid sterility. However, it has been shown how changes at the level of individual bases (slowly accumulating micromutations capable of impeding chromosomal pairing at meiosis, but not necessarily affecting the phenotype) could generate hybrid sterility (Forsdyke, 1996, 1998); such changes would not be seen when chromosomes were examined microscopically. Later-developing mechanisms of isolation, substituting for the primary micromutation-dependent mechanism, would include prezygotic isolating factors (e.g. mating preferences), and suddenly-appearing chromosomal macromutations. The latter are observed in chromosome studies of sterile hybrids (Chandley et al., 1974), and it is understandable that such obvious chromosomal rearrangements, probably occurring after the initial evolutionary divergence was complete, should have received attention as candidates contributing to the initial isolation process (White, 1978; King, 1993; Bush et al., 1977). Indeed, as Coyne and Orr (1998) point out, some types of macromutation are compatible with pairing. A strength of the micromutation proposal is that it allows fine degrees of compatibility, so that a rare reproductive variant would be more likely to find a partially matching physiological complement. The third criticism of Coyne and Orr (1998; Box 2), can be settled by the polyploidy test (Fig. 2; Winge, 1917), as was acknowledged by Theodosius Dobzhansky (1933, 1937). It is obvious that the pairing of homologs would require some gene products, and thus that mutations in these genes might impair chromosome pairing. Indeed, such genes might well be identified, mapped and successfully cloned (the fourth criticism), although Maside and Naviera (1996) have questioned the validity of some experiments claiming precise localization of genes having a major effect on hybrid sterility. Some applications of the polyploidy test by Dobzhansky (1933) do show a genic cause to apply in his particular cases of sterility. However, other applications of the test show that sterility often has a chromosomal basis (Darlington, 1932; Dobzhansky, 1937). It is incorrect to state that "This test has apparently not been repeated in any other hybridizations" (Coyne & Orr, 1998). As their fifth criticism, Coyne and Orr point to the sexual asymmetry of the incidence of hybrid problems in certain crosses. Their gene-centred viewpoint has dominated the debate on why sterility first affects the sex with different sex chromosomes (the heterogametic sex; Haldane, 1922), and |
| "forms an integral part of most hypotheses, either implicitly or explicitly" (Laurie, 1997). |
| These hypotheses must cope with "adaptive valleys" and other problems, but the lack of "wholly convincing explanations" has suggested that |
| "it seems possible that some simple explanation ... has eluded everyone" (Coyne, 1992). |
The alternative chromosomal viewpoint permits such an explanation. The antirecombinational effect of (C+G)% divergence as proposed for incipient speciation (Forsdyke, 1996), seems applicable in principle to the antirecombinational activity required to prevent recombination between genes required for incipient sexual differentiation (Forsdyke 1999b). As with speciation, subsequent macromutations (e.g. deletions in the Y chromosome) would replace the original (C+G)% divergence between genomic regions concerned with sexual differentiation, so that no trace of the original mechanism of divergence might be evident in many modern species. However, a head-start towards speciation would have been given to the heterogametic sex (the sex with X and Y gametes). Thus, to generate chromosomally-based hybrid sterility as part of the incipient speciation process, the homogametic sex would have to:
The heterogametic sex would have already completed much of the first of these. Regions of the two sex chromosomes would already be "reproductively isolated" from each other to allow differentiation of specific sexes.
Haldane's rule can also be manifest as hybrid inviability (Coyne, 1992). A proposed mechanism for the latter based on differences in dosage compensation (involving heteromorphic sex chromosomes; Forsdyke, 1995) would not be expected in to apply in crosses between "species" (varieties) of genera with homomorphic sex chromosomes. Mosquitos of the genus Aedes fit this criterion. Presgraves and Orr (1998) found for crosses between Aedes "species" that Haldane's rule was associated with hybrid sterility in the heterogametic (male) sex, not with hybrid inviability. |
Major potential "Modes of Speciation" (White, 1978) are "chromosomal" and "genic". Problems with the chromosomal mode may be resolved by taking into account differences in the "accent" of DNA, (C+G)%. This proposal is consistent with a variety of phenomena (e.g. the wide disparities in base composition between viruses with the potential to coinhabit a cell; Forsdyke, 1996), and is supported by bioinformatic analyses (Forsdyke, 1998). The present work traces the origins of the chromosomal hypothesis as an explanation for the progressive hybrid sterility by which incipient species are defined, and rebuts the case of Coyne and Orr (1998). It is shown elsewhere that subtle differences in base composition may also explain other intriguing properties of DNA (Bell et al., 1998; Bell & Forsdyke, 1999a,b). |
I thank J. Duffin, and G. Smith of Queen's University for comments on various aspects of this work, A. Barrett of Imperial College, London, for providing materials from the Huxley archive, and E. Barnes for a copy of her B.A. thesis on Romanes (Cambridge 1998).
REFERENCES
BATESON, W. (1904). Presidential address to the Zoological Section, British Association. In: William Bateson, F.R.S. Naturalist. His Essays and Addresses, (Bateson, B., ed). pp. 233-259. Cambridge: Cambridge Univ. Press.
BATESON, W. (1909a). Mendel's Principles of Heredity, pp. 72-73, 375. Cambridge: Cambridge University Press.
BATESON, W. (1909b). Heredity and variation in modern lights. In: William Bateson, F.R.S. Naturalist. His Essays and Addresses, (Bateson, B., ed). pp. 215-232. Cambridge: Cambridge Univ. Press.
BATESON, W. (1913). Problems of Genetics, pp. 119, 238-242. New Haven: Yale Univ. Press.
BATESON, W. (1922). Evolutionary faith and modern doubts. Science 55, 55-61.
BATESON, W. (1924). Progress in biology. In: William Bateson, F.R.S. Naturalist. His Essays and Addresses, (Bateson, B., ed). pp. 399-408. Cambridge: Cambridge Univ. Press.
BATESON, W. & SAUNDERS, E. R. (1902). Report I. In: Reports to the Evolution Committee of the Royal Society, pp. 1-160. London: Harrison.
BATESON, W., SAUNDERS, E. R. & PUNNETT, R. C. (1904). Report II. Experimental studies in the physiology of heredity. Reports to the Evolution Committee of the Royal Society, pp. 1-131. London: Harrison.
BELL. S. J., CHOW, Y. C., HO, J. Y. K. & FORSDYKE, D. R. (1998). Correlation of Chi orientation with transcription indicates a fundamental relationship between recombination and transcription. Gene 216, 285-292.
BELL, S. J. & FORSDYKE, D. R. (1999a). Accounting units in DNA. J. theor. Biol. 197, 51-61.
BELL, S. J. & FORSDYKE, D. R. (1999b). Deviations from Chargaff's second parity rule correlate with direction of transcription. J. theor. Biol. 197, 63-76.
BRONSON, E. C. & ANDERSON, J. N. (1994). Nucleotide composition as a driving force in the evolution of retroviruses. J. Mol. Evol. 38, 506-532.
BUSH, G. L., CASE, S. M., WILSON, A. C. & PATTON, J. L. (1977). Rapid speciation and chromosomal evolution in mammals. Proc. Natl. Acad. Sci. USA,74, 3942-3946.
CHANDLEY, A. C., JONES, R. C., DOTT, H. M., ALLEN, W. R. & SHORT, R. V. (1974). Meiosis in interspecific equine hybrids. 1. The male mule (Equus asinus X E. caballus) and hinny (E. caballus X E. asinus). Cytogen. Cell Genet. 13, 330-341.
CHARGAFF, E. (1951). Structure and function of nucleic acids as cell constituents. Fed. Proc. 10, 654-659.
CHEN, J-H., LE, S-Y., SHAPIRO, B., CURREY, K. M. & MAIZEL, J. V. (1990). A computational procedure for assessing the significance of RNA secondary structure. Comp. Applic. Biol. Sci. 6, 7-18.
COLEMAN, W. (1970). Bateson and chromosomes: conservative thought in science. Centaurus 15, 228-314.
COYNE, J. A. (1992). Genetics and speciation. Nature 355, 511-515.
COYNE, J. A. & ORR, H. A. (1998). The evolutionary genetics of speciation. Phil. Trans. R. Soc. B (London), 353, 287-305.
DARLINGTON, C. D. (1932). Recent Advances in Cytology, pp. 448-485. London: Churchill.
DARWIN, C. R. (1859). The Origin of Species by Means of Natural Selection or the Preservation of Favoured Races in the Struggle for Life. London: John Murray.
DARWIN, C. R. (1862). Notes on the causes of cross and hybrid sterility. In: The Correspondence of Charles Darwin, Vol. 10, 700-711, (Burkhardt, F., Porter, D. M., Harvey, J. & Topham, J. R., eds). Cambridge: Cambridge Univ. Press, 1997.
DELBOEUF, J. (1877). Les math�matiques et le transformisme. Une loi math�matique applicable � la th�orie du transformisme. La Revue Scientifique, 29, 669-679.
DOBZHANSKY, T. (1933). On the sterility of the interracial hybrids in Drosophila pseudoobscura. Genetics, 19, 397-403.
DOBZHANSKY, T. (1937). Genetics and the Origin of Species, pp. 288-291. New York: Columbia Univ. Press.
FORSDYKE, D. R. (1995). Fine tuning of intracellular protein concentrations, a collective protein function involved in aneuploid lethality, sex-determination, and speciation. J. theor. Biol. 172, 335-345.
FORSDYKE, D. R. (1996). Different biological species "broadcast" their DNAs at different (G+C)% "wavelengths". J. theor. Biol. 178, 405-417.
FORSDYKE, D. R. (1998). An alternative way of thinking about stem-loops in DNA: a case study of the G0S2 gene. J. theor. Biol. 192, 489-504.
FORSDYKE, D. R. (1999a). The origin of species, revisited. Queen's Quarterly 106, 112- 133.
FORSDYKE, D. R. (1999b). Haldane's rule exists since reproductive and species differentiations both require reproductive isolation. FASEB. J. 13, A1543.
GOULD, S. J. (1980). Is a new and general theory of evolution emerging? Paleobiology 6, 119-130.
GULICK, A. (1932). John Thomas Gulick: Evolutionist and Missionary, pp. 497-499. Chicago: Univ. Chicago Press.
GULICK, J. T. (1872). On diversity of evolution under one set of external conditions. J. Linn. Soc. (Zool.), 11, 496-505.
GUYER, M. F. (1902). Hybridism and the germ cell. Bull. Univ. Cincinnati 21, 1-20.
HALDANE, J. B. S. (1922). Sex ratio and unisexual sterility in hybrid animals. J. Genet. 12, 101-109.
HALDANE, J. B. S. (1957). The theory of evolution before and after Bateson. J. Genet. 56, 11-27.
HOOKER, J. (1862). Letter to Bates. In: The Correspondence of Charles Darwin, Vol. 10, 127-130. (Burkhardt, F., Porter, D. M., Harvey, J. & Topham, J. R., eds). Cambridge: Cambridge Univ. Press, 1997.
HUNTER, N., CHAMBERS, S. R., LOUIS, E. J. & BORTS, R. H. (1996). The mismatch repair system contributes to meiotic sterility in an interspecific yeast hybrid. Eur. Mol. Biol. Org. J. 15. 1726-1733.
HUXLEY, T. H. (1863). Six Lectures to Working Men. In: Darwiniana. Collected Essays, pp. 463-466. London: Macmillan, 1893.
KING, M. (1993). Species Evolution. The Role of Chromosome Change. Cambridge: Cambridge Univ. Press.
KLECKNER, N. (1997). Interactions between and along chromosomes during meiosis. Harvey Lect. 91, 21-45.
LAURIE, C. C. (1997). The weaker sex is heterogametic. 75 years of Haldane's rule. Genetics 147, 937-951.
LESCH, J. E. (1975). The role of isolation in evolution: George J. Romanes and John T. Gulick. Isis 66, 483-503.
MASIDE, X. R. & NAVEIRA, H. F. (1996). A polygenic basis of hybrid sterility may give rise to spurious localizations of sterility factors. Heredity 77, 488-492.
MATASSI, G., MELIS, R., MACAYA, G. & BERNARDI, G. (1991). Compositional bimodality of the nuclear genome of tobacco. Nucleic Acids Res. 19, 5561-5567.
OLBY, R. (1985). The Origins of Mendelism, 2nd edition. Chicago: Univ. Chicago Press.
ORR, H. A. (1996). Dobzhansky, Bateson, and the genetics of speciation. Genetics 144, 1331-1335.
PAGE, A. W. & ORR-WEAVER, T. L. (1996) Stopping and starting the meiotic cycle. Curr. Opin. Genet. Devel. 7, 23-31.
PRESGRAVES, D. C. & ORR, H. A. (1998). Haldane's rule in taxa lacking a hemizygous X. Science 282, 952-954.
PROVINE, W. B. (1986). Sewell Wright and Evolutionary Biology, p. 207. Chicago: Univ. Chicago Press.
ROCCO, V. & NICOLAS, A. (1996). Sensing of DNA non-homology lowers the initiation of meiotic recombination in yeast. Genes to Cells 1, 645-661.
ROMANES, G. J. (1886). Physiological selection. An additional suggestion on the origin of species. J. Linn. Soc. (Zool.) 19, 337-411.
ROMANES, G. J. (1887). Physiological Selection. Nineteenth Century 21, 59-80.
ROMANES, G. J. (1897). Darwin, and After Darwin. Volume 3. Isolation and Physiological Selection. London: Longmans and Green.
SCHACHTEL, G. A., BUCHER, P., MOCARSKI, E. S., BLAISDELL, B. E. & KARLIN, S. (1991). Evidence for selective evolution in codon usage in conserved amino acid segments of human alphaherpesvirus proteins. J. Mol. Evol. 33, 483-494.
SHAW, G. B. (1913). Pygmalion. In: Bernard Shaw. Complete Plays with Prefaces. Vol. 1, 190-295. New York: Dodd, Mead & Co., 1963.
SUEOKA, N. (1961). Compositional correlation between deoxyribonucleic acid and protein. Cold Spring Harbor Symp. Quant. Biol. 26, 35-43.
SUTTON, W. S. (1903). The chromosomes in heredity. Biol. Bull. 4, 231-251.
WALLACE, A. R. (1886). Physiological selection and the origin of species. Nature 34, 467-468.
WHITE, M. J. D. (1978). Modes of Speciation, pp. 323-349. San Francisco: Freeman.
WINGE, �. (1917). The chromosomes, their number and general importance. Comp. Rend. Trav. Carls. Lab. 13, 131-275.
WONG, B. C., CHIU, S-K., & CHOW, S. A. (1998). The role of negative superhelicity and length of homology in the formation of paranemic joints promoted by RecA protein. J. Biol. Chem. 273, 12120-12127.
WYATT, G. R. (1952). The nucleic acids of some insect viruses. J. Gen. Phys. 36, 201- 289.
YIN, P. D. & HU, W-S. (1997). RNAs from genetically distinct retroviruses can copackage and exchange genetic information. J. Virol. 71, 6237-6242.
![]()
|
End Note July 2001: Regarding section 10 above (i.e. metaphors for the exclusionary role of secondary information), it is of interest that William Bateson's son Gregory (1904-1978; named after Gregor Mendel) who was an anthropologist/psychologist became interested in the hierarchy of information levels involved in human communication. He and his colleagues noted (1956) that schizophrenic patients often appeared unable to recognize the "context" in which primary information is given:
More simply stated, there is no smile or wink (secondary information) acting as a "classifying metamessage." His biographer, David Lipsett writes (1980; p. 208):
End Note February 2002: In a paper entitled "Dual information in DNA and the evolution of the genetic code," Tamar Schaap (1971, J. Theor. Biol. 32, 293-98; Click Here) pointed out that a sequence encoding a protein sequence could also contain information for recognition by proteins involved in DNA transactions (recombination, replication, transcription, repair). Thus, if a mutation did not change an amino acid it was not, by default, "neutral." This lone voice was of little avail against the neutralist juggernaut. Believing in a superfluity of information space in genomes, the neutralists dismissed mutations with no obvious phenotypic effect, and genome space with no obvious phenotypic role, as "neutral" or "junk," respectively. End Note January 2007: In his book Mind and Nature (1979) Gregory Bateson referred to higher hierarchical levels (i.e. secondary information) as "typological" that contrasted with the "synthetic" views of Mayr, who favoured the so-called "modern synthesis" of Darwinian and Mendelian ideas (i.e. primary information):
Bateson also pointed to possible conflicts between different levels of information (primary, secondary, etc.):
|
![]()
Return to: HomePage (Click Here)
Return to: Evolution Index (Click Here)
Return to: Mind page (Click Here)
Go to: Haldane's Rule (Click Here)
Go to: Bateson & Saunders, 1902 (Click Here)
Go to: Bateson's Addresses (Click Here)
Go to: Opposition to Bateson (Click Here)
Go to: Bateson and the "residue" (Click Here)
Go to: Opposition to above paper (Click Here)
Go to: Reply to above opposition (Click Here)
![]()
This page was established circa 1999 and last edited on 12 September 2010 by Donald Forsdyke


 This would predict that
incipient speciation, manifest as some degree of hybrid sterility
when "varieties" were crossed, would be
detected at the earliest stage in the heterogametic sex (i.e. Haldane's rule), even in
genera with homomorphic sex
chromosomes (i.e. when X and Y chromosomes appear
identical). Later, autosomal differentiation would increase, and reproductive isolation
between the "varieties" (then, by definition,
"species") would be complete.
This would predict that
incipient speciation, manifest as some degree of hybrid sterility
when "varieties" were crossed, would be
detected at the earliest stage in the heterogametic sex (i.e. Haldane's rule), even in
genera with homomorphic sex
chromosomes (i.e. when X and Y chromosomes appear
identical). Later, autosomal differentiation would increase, and reproductive isolation
between the "varieties" (then, by definition,
"species") would be complete.